
 >
>微量元素及低温对生物膜硝化性能的影响
时间:
为探究微量元素及低温下环境条件改变对生物膜反应器硝化性能的影响,以含(NH_{4}^{+}-N)模拟废水为处理对象,研究了微量元素、低温、曝气量和流速对生物膜反应器硝化性能的影响,利用16S rRNA高通量测序技术分析微生物群落结构。结果表明,微量元素显著影响硝化性能((P<0.0001)),进水中添加微量元素后(NH_{4}^{+}-N)去除负荷从(0.93 kg/(m^{3}·d))增加至(1.63 kg/(m^{3}·d)),(NO_{3}^{-}-N)生成负荷从(0.23 kg/(m^{3}·d))提高至(1.21 kg/(m^{3}·d))。低温会对硝化细菌产生影响,其中亚硝酸盐氧化菌(NOB)对低温冲击敏感,氨氧化菌(AOB)则对低温有抗冲击能力。曝气量降低导致反应器内溶解氧(DO)不足,进而影响硝化性能。流速改变没有对硝化性能产生显著影响。微生物群落结构分析结果表明,硝化细菌Nitrosomonas和Nitrospira在运行过程中富集,保证了低温条件下反应器仍能具有稳定的硝化性能。
关键词: 硝化反应; 生物膜; 微量元素; 低温; 曝气量; 流速
论文《微量元素及低温对生物膜硝化性能的影响》发表在《中国环境科学》,版权归《中国环境科学》所有。本文来自网络平台,仅供参考。
与传统活性污泥法相比,生物膜法具有处理负荷高、抗冲击能力强、污泥产量少等优点,越来越得到研究人员关注[1-3]。围绕硝化生物膜技术已开展了大量研究,包括探究填料类型、进水负荷、水力停留时间(HRT)等对硝化性能的影响[4-6],以及硝化生物膜中微生物的组成及分布情况[7-8]。但如何提高系统的稳定性和效率仍有待研究。
环境条件会影响微生物活性、污水处理效率和出水水质[9]。已有研究者开展了盐度、碳氮比(C/N)、溶解氧(DO)以及温度对生物膜反应器脱氮性能的影响研究。研究发现,在高盐度下生物膜的反硝化能力受抑制程度远比硝化能力小,快速提升盐度会对生物膜脱氢酶活性造成抑制[10]。进水C/N提高将导致异养菌和硝化菌竞争生物膜表面空间和DO,抑制硝化菌增殖的同时硝化活性也会迅速降低[11]。反应器内DO浓度越高,生物膜中好氧区所占比例越大,但DO传递阻力也随之增大[12]。低温条件下生物膜中硝化细胞拷贝数会小幅上升,同时伴随着生物量的增加,能从一定程度上弥补低温下硝化活性的衰减[13]。但是,目前关于环境条件对硝化生物膜反应器性能影响的研究仍不充足,尤其是反应器长期运行下制约硝化性能的重要环境条件。
因此,本文以含(NH_{4}^{+}-N)模拟废水为处理对象,建立硝化生物膜反应器,探究微量元素和低温冲击对硝化生物膜反应器性能的影响,以及曝气量和流速对低温硝化生物膜反应器性能的影响;考察生物膜胞外聚合物(EPS)含量及成分变化情况,分析生物膜中微生物群落结构变化,确定优势菌群并对氮代谢功能基因进行预测,为实现不同环境条件下硝化生物膜反应器的稳定长期运行提供参考。
1 材料与方法
1.1 实验装置
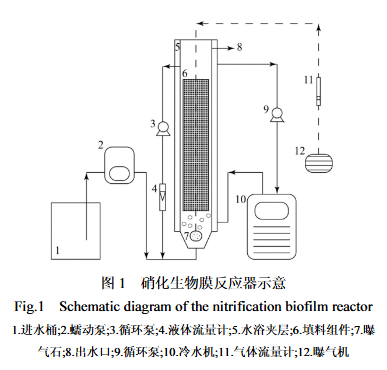
反应器有效工作体积为5.15L,反应器内装有体积为1.62L的网状填料组件,材质为聚丙烯。反应器底部设有曝气石,通过曝气机向反应器内提供DO。反应器外设有水浴夹层,可通过冷水机(广东海利,HC-300A)和循环泵(上海新西山,MP-6R)实现硝化生物膜反应器低温运行。
图1 硝化生物膜反应器示意 Fig.1 Schematic diagram of the nitrification biofilm reactor
(注:1.进水桶;2.蠕动泵;3.循环泵;4.液体流量计;5.水浴夹层;6.填料组件;7.曝气石;8.出水口;9.循环泵;10.冷水机;11.气体流量计;12.曝气机)
1.2 实验用水与接种污泥
实验用水为模拟废水,以(NH_{4}Cl)作为(NH_{4}^{+}-N)来源,(NaH_{2}PO_{4})作为(PO_{4}^{3-}-P)来源,(NaHCO_{3})提供硝化过程所需碱度,添加量按每去除1.00g (NH_{4}^{+}-N)消耗7.14g碱度计算。硝化生物膜反应器运行第0~16d进水中未添加微量元素,待反应器性能稳定,第17d开始向进水中加入1mL/L微量元素溶液,其组成为(mg/L):(MgSO_{4}·7H_{2}O) 200、(KH_{2}PO_{4}) 300、(CaCl_{2}·2H_{2}O) 100、(FeCl_{2}·6H_{2}O) 100、(MnCl_{2}·4H_{2}O) 50、(CoCl_{2}·6H_{2}O) 30、(NiCl_{2}·6H_{2}O) 30、(CuCl_{2}·6H_{2}O) 20、(ZnCl_{2}·6H_{2}O) 70、(NaMoO_{4}·6H_{2}O) 30、(H_{3}BO_{3}) 60。模拟废水水质指标如表1所示。
表1 模拟废水水质 Table 1 Quality of the synthetic wastewater
| (NH_{4}^{+}-N) (mg/L) | (NO_{2}^{-}-N) (mg/L) | (NO_{3}^{-}-N) (mg/L) | (PO_{4}^{3-}-P) (mg/L) | (CaCO_{3})碱度 (mg/L) | pH值 |
| 53.30±9.70 | 2.21±1.93 | 2.32±1.36 | 1.87±0.22 | 444.82±25.66 | 7.84±0.19 |
接种污泥取自河南省郑州市五龙口污水处理厂二沉池,混合液悬浮固体浓度(MLSS)为3850mg/L。反应器启动时先将接种污泥加入反应器中,开启循环泵使填料组件与接种污泥充分混合,将曝气量设定为0.8L/min闷曝3d,排出呈悬浮状态的接种污泥,之后反应器开始连续进水运行。
1.3 实验运行方案
实验共分为2个运行阶段,运行阶段I为中温运行阶段,探究微量元素对硝化生物膜反应器性能的影响以及反应器长期运行效果;阶段II为低温运行阶段,探究低温冲击、曝气量和流速变化对硝化生物膜反应器性能的影响。运行方案如表2所示。
表2 实验运行方案 Table 2 Experimental operation scheme
| 运行阶段 | 时间 (d) | 曝气量 (L/min) | HRT (h) | 水温 (℃) | 流速 (m/h) | 微量元素 |
| I | 0~16 | 0.8 | 3.0~10.0 | 23~25 | 10 | 未添加 |
| | 17~107 | 0.8 | 2.0~4.0 | 23~29 | 10 | 添加 |
| II | 108~146 | 0.8 | 3.5~5.0 | 10~18 | 10 | 添加 |
| | 147~183 | 0.2~0.7 | 5.0 | 12~16 | 10 | 添加 |
| | 184~338 | 0.2 | 4.5~5.0 | 10~21 | 12~55 | 添加 |
1.4 分析项目与检测方法
(NH_{4}^{+}-N)采用纳氏试剂光度法测定,(NO_{2}^{-}-N)采用N-(1-萘基)-乙二胺光度法测定,(NO_{3}^{-}-N)采用紫外分光光度计法测定,(PO_{4}^{3-}-P)采用钼锑抗分光光度法测定,碱度采用酸碱指示剂滴定法测定,MLSS和MLVSS采用重量法测定[14],pH值使用便携式pH计(上海雷磁,PHB-4)测定,DO使用便携式溶解氧分析仪(美国哈希,HQ30d)测定。
对接种污泥以及反应器低温运行达到稳定时(第267d和第330d)的生物膜样本进行收集,测定EPS含量并进行16S rRNA高通量测序分析。EPS采用热提取法进行提取[15],其中多糖(PS)采用苯酚-硫酸法测定,蛋白质(PN)采用修正的Lowry法测定[16]。微生物样本交由生工生物工程(上海)股份有限公司进行16S rRNA高通量测序分析。使用E.Z.N.A™ Mag-Bind Soil DNA Kit(OMEGA)试剂盒提取DNA,使用Qubit 4.0(Thermo, USA)测定DNA浓度,合格后对细菌16S rRNA基因的V3~V4区进行PCR扩增,引物为341F(CCTACGGGNGGCWGCAG)和805R(GACTACHVGGGTATCTAATCC),之后使用Illumina MiSeq系统进行高通量测序。
绘制操作分类单元(OTUs)稀疏曲线,用Mothur软件(3.8.31版)计算α多样性指数。使用PICRUSt软件(1.1.4版)对细菌进行功能预测分析,通过将现有的16S rRNA基因测序数据与已知代谢功能的微生物参考基因组数据库进行比较,预测细菌的代谢功能。使用GraphPad Prism软件(9.5版)对不同环境条件下(NH_{4}^{+}-N)去除负荷、(NO_{2}^{-}-N)生成负荷、(NO_{3}^{-}-N)生成负荷和总无机氮(TIN)去除率相关数据进行One-Way ANOVA分析和t检验分析。
1.5 计算方法
(NH_{4}^{+}-N)去除负荷((L_{NH_{4}^{+}-N}),(kg/(m^{3}·d)))、(NO_{2}^{-}-N)生成负荷((L_{NO_{2}^{-}-N}),(kg/(m^{3}·d)))、(NO_{3}^{-}-N)生成负荷((L_{NO_{3}^{-}-N}),(kg/(m^{3}·d)))计算方法分别如式(1)、(2)、(3)所示:
式中:(NH_{4}^{+}-N_{Inf.})为进水(NH_{4}^{+}-N)浓度(mg/L);(NH_{4}^{+}-N_{Eff.})为出水(NH_{4}^{+}-N)浓度(mg/L);(NO_{2}^{-}-N_{Inf.})为进水(NO_{2}^{-}-N)浓度(mg/L);(NO_{2}^{-}-N_{Eff.})为出水(NO_{2}^{-}-N)浓度(mg/L);(NO_{3}^{-}-N_{Inf.})为进水(NO_{3}^{-}-N)浓度(mg/L);(NO_{3}^{-}-N_{Eff.})为出水(NO_{3}^{-}-N)浓度(mg/L);HRT为水力停留时间(h);(V_{1})为反应器有效工作体积(L);(V_{2})为填料组件体积(L)。
2 结果与讨论
2.1 微量元素对硝化生物膜反应器性能的影响
如图2所示,反应器运行至第3d出水(NH_{4}^{+}-N)浓度可降至1mg/L以下,(NH_{4}^{+}-N)去除率可达到98.67%,(NH_{4}^{+}-N)去除负荷为0.43(kg/(m^{3}·d))。之后HRT逐渐从10h缩短至3h,虽然(NH_{4}^{+}-N)去除率从91.62%下降至68.27%,但(NH_{4}^{+}-N)去除负荷逐渐从0.20(kg/(m^{3}·d))提高至1.10(kg/(m^{3}·d))。此时出水中氮氧化物((NO_{x}^{-}-N))主要以(NO_{2}^{-}-N)形式存在,且浓度持续增加至32.08mg/L。通过理论计算可知(图2(d)),(NO_{3}^{-}-N)生成负荷始终维持在较低水平(0.15~0.38(kg/(m^{3}·d))),推测造成这一现象的原因可能是反应器启动初期生物膜内亚硝酸盐氧化菌(NOB)数量不足或活性不高,导致(NO_{2}^{-}-N)在缩短HRT提负荷的过程中不断积累。
微生物生长需要各种营养物质,而微量元素在微生物生长过程中扮演重要角色,因为它们在酶的组成、电子传递及代谢过程中发挥关键作用[17]。有研究表明铁、硼、锰的添加会使氨氮去除速率提高[18],铜的添加使氨单加氧酶的活性大大增加[19]。因此在硝化生物膜反应器运行至第17d时开始向进水中添加微量元素溶液。从图2(a)和图2(d)可以看出,微量元素添加后5d内(NH_{4}^{+}-N)去除率从68.27%提高至82.97%,(NO_{3}^{-}-N)生成速率从0.23(kg/(m^{3}·d))提高至0.68(kg/(m^{3}·d))。之后随着反应器运行,(NH_{4}^{+}-N)去除负荷增加至1.63(kg/(m^{3}·d)),(NO_{3}^{-}-N)生成负荷逐渐提高至1.21(kg/(m^{3}·d)),出水中的(NO_{2}^{-}-N)浓度逐渐下降至5.26mg/L。结合统计学分析结果(图3(a)),微量元素添加显著促进生物膜反应器内硝化速率提高((P<0.0001)),由此推测微量元素对生物膜系统中NOB的富集和活性表达起到至关重要的作用,可快速提高(NH_{4}^{+}-N)去除速率以及调控出水中(NO_{x}^{-}-N)存在形式。
(a) (NH_{4}^{+}-N)浓度和去除率 (b) 出水(NO_{2}^{-}-N)浓度和水温
(c) (NH_{4}^{+}-N)去除负荷和HRT (d) (NO_{2}^{-}-N)、(NO_{3}^{-}-N)生成负荷和流速
图2 硝化生物膜反应器运行期间脱氮性能及运行参数变化 Fig.2 Nitrogen removal performance and operating parameters during the operation of nitrification biofilm reactor
(a) 微量元素对负荷的影响 (b) 温度对负荷的影响
(c) 曝气量对负荷的影响 (d) 曝气量对TIN去除率的影响
图3 运行参数与反应器硝化性能统计学分析 Fig.3 Statistical analysis of operating parameters and nitrification performance of the biofilm reactor
(注:****(P<0.0001),***(P<0.001))
反应器运行至第48d时,肉眼观察到生物膜状态变差,有少部分生物膜从填料表面脱落,堆积在填料组件底部,(NH_{4}^{+}-N)去除负荷和(NO_{3}^{-}-N)生成负荷明显下降。此时将HRT从2h延长至2.5h,反应器处理效果未改善。值得注意的是,在观察到生物膜状态变差之前(运行第31d)已经检测到(NH_{4}^{+}-N)去除负荷逐渐从1.63(kg/(m^{3}·d))下降至1.32(kg/(m^{3}·d)),(NO_{3}^{-}-N)生成负荷从1.21(kg/(m^{3}·d))下降至0.82(kg/(m^{3}·d)),在观察到生物膜形状变差后(NH_{4}^{+}-N)去除负荷又持续下降至0.90(kg/(m^{3}·d))。因此,水质指标能及时响应反应器内生物膜的性能变化。反应器运行至第72d,通过排泥去除填料组件底部堆积的微生物,(NH_{4}^{+}-N)去除负荷和(NO_{3}^{-}-N)生成负荷逐渐恢复至1.20~1.42(kg/(m^{3}·d))和0.94~1.38(kg/(m^{3}·d)),堆积的生物膜可能影响了反应器内部物质传质,导致硝化效果下降。
为了开展后续低温实验,第97d在反应器外围缠绕水浴夹层管,这一过程导致反应器发生震动,部分生物膜脱落,对硝化性能产生不利影响,(NH_{4}^{+}-N)去除负荷从1.38(kg/(m^{3}·d))降至0.84(kg/(m^{3}·d)),(NO_{3}^{-}-N)生成负荷从1.45(kg/(m^{3}·d))降至1.06(kg/(m^{3}·d))。该现象说明硝化菌在填料表面附着不牢固,易受外界冲击而流失。
2.2 低温冲击对硝化生物膜反应器性能的影响
低温环境下微生物的活性降低,生长速度减慢,导致生物处理系统对污染物的去除效率下降[20]。本实验中降温分为两个阶段,第一阶段为108~129d,水温以每天0.5~2.0℃从29℃逐渐降低至15℃,该过程明显观察到(NO_{3}^{-}-N)生成负荷从0.87(kg/(m^{3}·d))降低至0.37(kg/(m^{3}·d))((P<0.001)),而出水(NO_{2}^{-}-N)逐渐积累(从2.47mg/L增加至15.16mg/L),而(NH_{4}^{+}-N)去除负荷始终维持在0.80~0.91(kg/(m^{3}·d))。该实验现象表明低温首先影响NOB活性,导致出水(NO_{2}^{-}-N)积累,而AOB则对低温有一定的抗冲击能力。杨思敏等的研究结果表明,水温降低会导致(NH_{4}^{+}-N)去除负荷和(NO_{3}^{-}-N)生成负荷同时降低,这可能是因为该研究将水温降至更低水平(10和5℃),AOB的活性也受到抑制[21]。
第129~132d,由于冷却循环管路堵塞,反应器内水温恢复至25℃(期间未检测进出水指标)。第133d测得(NO_{3}^{-}-N)生成负荷恢复至0.75(kg/(m^{3}·d)),(NH_{4}^{+}-N)去除负荷保持在0.86(kg/(m^{3}·d))。随后开始第二阶段降温,一天内将水温从25℃降至12.5℃,(NH_{4}^{+}-N)去除负荷和(NO_{3}^{-}-N)生成负荷没有显著变化,分别为0.91和0.71(kg/(m^{3}·d))。之后随着反应器运行,硝化微生物逐渐适应低温环境,(NH_{4}^{+}-N)去除负荷逐渐提高至1.10(kg/(m^{3}·d)),(NO_{3}^{-}-N)生成负荷逐渐提高至0.91(kg/(m^{3}·d))。
在第一阶段降温期间,观察到生物膜表面产生少量白色粘性物质,推测为微生物为对抗低温分泌的EPS,因此第二次快速降温时反应器性能没有明显降低。
2.3 曝气量对低温硝化生物膜反应器性能的影响
为探究曝气量对低温硝化生物膜反应器性能的影响,从第148d开始逐渐将曝气量从0.8L/min调至0.2L/min。曝气量从0.8L/min下调至0.3L/min的过程中,反应器内DO逐渐从7.92mg/L降至6.70mg/L,反应器性能没有受到显著影响(图3(c)),(NH_{4}^{+}-N)去除负荷维持在0.82~1.01(kg/(m^{3}·d)),(NO_{3}^{-}-N)生成负荷维持在0.75~0.93(kg/(m^{3}·d))。当曝气量降至0.2L/min时,出水DO降至5.5mg/L以下,此时反应器性能显著变差,(NH_{4}^{+}-N)去除负荷为0.60~0.96(kg/(m^{3}·d)),(NO_{3}^{-}-N)生成负荷为0.46~0.76(kg/(m^{3}·d))。此外,曝气量降低还显著影响了TIN的去除率(图3(d)),当曝气量为0.3~0.8L/min时,平均TIN去除率为7.21%,当曝气量降至0.2L/min时,平均TIN去除率为15.44%。曝气量的降低一方面会影响硝化菌活性,另一方面可能会降低生物膜内部DO浓度,从而促进反硝化菌生长富集,在生物膜内形成同步硝化反硝化生物脱氮路径,造成TIN损失[22-23]。
2.4 流速对低温硝化生物膜反应器性能的影响
反应器内流速与传质、生物膜更新等密切相关,是影响反应器性能的重要运行参数[24]。第185~282d,流速逐渐从10m/h提高至55m/h,第283~338d,流速逐渐从55m/h下调至10m/h。每次改变流速后会出现负荷波动的现象,可能是流速变化对微生物造成冲击,但随着反应器运行,负荷可以逐渐恢复稳定。但从整体上来看,流速改变没有对硝化性能造成显著的影响。在低曝气量下(0.2L/min)明显观察到气泡对水体的扰动减弱,虽然期望通过提高流速来增加传质效果,但气泡的扰动作用似乎要强于流速。
2.5 微生物分析
2.5.1 EPS组成分析
如图4所示,接种污泥以及生物膜样本中EPS的主要成分是PN,这与大多数研究结果一致[25-26]。与接种污泥相比,生物膜样本中含有更多PN和PS。研究表明,EPS是影响生物膜形成的主要因素,PS提供支撑骨架,PN应对环境冲击[27]。第330d生物膜样本与第267d相比PS含量略有降低,导致PN/PS升高。有研究表明PN/PS越大,生物膜与填料结合越紧密、厚实和稳定[28],可能是流速改变引起的水力剪切力变化,使生物膜结构变得紧密。
图4 微生物样本中PS和PN含量及PN/PS Fig.4 Contents of PS and PN in microbial samples and PN/PS
2.5.2 微生物群落多样性分析
从表3可知,接种污泥的Ace指数、Chao指数和Shannon指数均大于生物膜样本,且Simpson指数小于生物膜样本,表明生物膜样本中微生物群落丰度和多样性低于接种污泥。这主要是由于絮状污泥和生物膜的生长环境存在差异,生物膜内的微生物生长空间有限,物质的传递受膜扩散影响,这就使得竞争能力较差的微生物逐渐被淘汰[29]。接种污泥样本的Shannoneven指数较高,表明微生物群落均匀度高,物种间分布均匀,而生物膜样本群落均匀度较低,主要与生物膜中底物和氧气供应存在浓度梯度有关,导致微生物群落分布不均。
表3 微生物样本α多样性指数 Table 3 The α diversity index of microbial samples
| 样本 | Ace | Chao | Shannon | Simpson | Shannoneven |
| 接种污泥 | 2640.37 | 2547.79 | 6.27 | 0.01 | 0.80 |
| 第267d | 1730.41 | 1616.17 | 4.71 | 0.03 | 0.65 |
| 第330d | 1567.53 | 1506.71 | 4.64 | 0.03 | 0.64 |
由图5可见,接种污泥中的OTUs总数最多,共有2499个,运行第267d和第330d的生物膜样本中OTUs总数低于接种污泥。3个微生物样本的共有OTUs为509个。接种污泥中的独有OTUs最多,为1346个,表明接种污泥中生物多样性最高,而第267d和第330d独有OTUs仅有465个和400个,表明在反应器运行过程中由于环境的变化,某些微生物逐渐被淘汰,导致OTUs数减少。
图5 微生物样本OTUs的Upset图 Fig.5 Upset plot of the OTUs for microbial samples
2.5.3 微生物群落优势菌分析
如图6(a)所示,3个微生物样本中的优势菌门为Bacteroidota、Proteobacteria和Chloroflexi,它们是污水处理系统中常见的优势菌门。Bacteroidota对生物脱氮有重要作用[30],其相对丰度从接种污泥中的18.31%逐渐增加至第330d的31.55%。Proteobacteria中包含了多种功能类型的微生物,如有机物降解、脱氮、除磷,在污水处理过程中发挥积极作用[31-32]。Chloroflexi的相对丰度从接种污泥中的15.91%持续下降至第330d的7.99%,有研究称其具有反硝化功能[33]。与硝化反应密切相关的Nitrospirota在接种污泥中的相对丰度为1.10%,随着反应器运行其相对丰度逐渐提高至第267d的3.36%和第330d的5.39%,硝化菌的富集有助于反应器在多变的环境条件下仍能维持较稳定的硝化性能。此外还观察到Acidobacteriota的相对丰度从接种污泥中的5.00%逐渐增加至第330d的16.17%,有研究称Acidobacteriota中的部分功能菌具有还原氮氧化物的能力[34],表明反应器内充足的(NO_{3}^{-}-N)促进其不断富集。
图6(b)为微生物样本中相对丰度前10以及与脱氮相关的功能菌。Nitrosomonas是AOB,功能是将(NH_{4}^{+}-N)氧化为(NO_{2}^{-}-N),其相对丰度从接种污泥中的0.20%逐渐增加至第330d的0.66%。Nitrospira是NOB,进一步将(NO_{2}^{-}-N)氧化为(NO_{3}^{-}-N)来实现反应器内的全程硝化,其相对丰度从接种污泥中的1.10%逐渐增加至第330d的5.39%。此外,生物膜样本中还存在另一种新型NOB,有研究表明Candidatus_Nitrotoga在低温条件下也能发挥良好的硝化作用[35]。Candidatus_Nitrotoga在接种污泥中的相对丰度<0.01%,在反应器运行过程中相对丰度略有富集,第267d和第330d相对丰度为0.03%和0.01%,表明反应器内低温环境有利于Candidatus_Nitrotoga富集。生物样本中还检测到大量反硝化菌norank_Saprospiraceae、norank_SJA-28、UTCFX1、Defluviimonas、OLB17、Hirschia和Terrimonas,其中UTCFX1可在低温条件下进行反硝化[36]。此外,在生物膜样本中还观察到厌氧氨氧化菌Candidatus_Kuenenia富集,其在接种污泥中的相对丰度<0.01%,在第267d为0.03%和第330d为0.01%,表明在生物膜内部可能形成厌氧环境,(NH_{4}^{+}-N)在氧化为(NO_{2}^{-}-N)时被厌氧氨氧化菌利用直接转化为(N_{2})。生物膜内反硝化菌和厌氧氨氧化菌的存在是导致反应器内TIN损失的主要原因。norank_Candidatus_Pacebacteria可能也是一种潜在的脱氮菌,其在同步硝化反硝化反应器是优势菌。norank_Pedosphaeraceae也是生物膜样本中的优势菌属,主要与EPS分泌有关。
(a) 门水平 (b) 属水平
图6 微生物群落结构组成分析 Fig.6 Analysis of the composition of microbial community structure
2.5.4 氮代谢功能基因分析
利用KEGG数据库对微生物的氮代谢途径相关功能基因进行分析。如图7所示,所有微生物样本均存在5种氮代谢途径,分别为硝化、反硝化、异化硝酸盐还原、同化硝酸盐还原和固氮,氮代谢功能基因有26个。硝化过程相关的基因包括氨单加氧酶基因pmoABC-amoABC、羟胺氧化酶基因hao及亚硝酸盐氧化还原酶基因nxrAB。第267d hao和pmoABC-amoABC相对丰度与接种污泥相比下降,但此时反应器仍具有一定硝化功能,氨氮去除负荷为0.48(kg/(m^{3}·d))。pmoABC-amoABC和hao相对丰度总和小于nxrAB,表明亚硝酸盐向硝酸盐的转化作用在硝化过程中占据主导地位,这也是反应器出水主要以(NO_{3}^{-}-N)形式存在的原因。反应器内涉及反硝化、异化硝酸盐还原、同化硝酸盐还原和固氮的基因始终占据一定丰度,表明反应器内存在多种氮代谢路径,这也与反应器出水存在少量TIN损失相吻合。由于KEGG注释的局限性,与厌氧氨氧化氮代谢途径至关重要的联氨合成酶基因Hzs和联氨脱氢酶基因Hdh未被检测到[37]。
图7 氮代谢功能基因的相对丰度 Fig.7 Relative abundance of the nitrogen metabolism functional genes
3 结论
3.1 微量元素是影响硝化生物膜反应器性能的关键因素。进水中添加微量元素后(NH_{4}^{+}-N)去除负荷和(NO_{3}^{-}-N)生成负荷可快速显著提高((P<0.0001))。
3.2 水温降低会对硝化生物膜反应器性能产生不利影响。低温冲击会影响NOB活性,导致出水(NO_{2}^{-}-N)浓度逐渐从2.47mg/L积累至15.16mg/L,而AOB则对低温有一定的抗冲击能力,(NH_{4}^{+}-N)去除负荷始终维持在0.80~0.91(kg/(m^{3}·d))。
3.3 曝气量降低会导致反应器内DO不足,对硝化性能和TIN去除率产生影响。当曝气量降至0.2L/min时,硝化性能显著下降((P<0.001)),平均TIN去除率从7.21%提高至15.44%。
3.4 低温条件下流速的变化没有对硝化性能造成显著影响。
3.5 与硝化过程密切相关的Nitrosomonas(AOB)和Nitrospira(NOB)随着反应器运行逐渐富集,保证了反应器在低温条件下仍能具有较好的硝化性能。
3.6 氮代谢功能基因分析结果表明反应器内存在硝化、反硝化、异化硝酸盐还原、同化硝酸盐还原和固氮多种氮代谢路径,这与反应器出水存在少量TIN损失相吻合。
参考文献
[1] 敬双怡, 宋子洋, 刘超, 等. MBBR工艺中SNEDPR的启动及性能研究[J]. 中国环境科学, 2022,42(7):3121-3129.
[2] Huang Y N, Hu T Q, Lin B C, et al. Microplastics-biofilm interactions in biofilm-based wastewater treatment processes: A review[J]. Environmental Pollution, 2024,361:124836.
[3] Zhang X N, Al-Dhabi N A, Zhu Z X, et al. Robust PD/A biofilms sustain advanced real municipal wastewater treatment: Reinforcement of interspecific communication and electron transfer[J]. Chemical Engineering Journal, 2024,499:156085.
[4] 钟镭, 王盟, 赵白航, 等. 载体对短程硝化生物膜特性的影响及机理探究[J]. 工业水处理, 2021,41(2):75-79.
[5] 王荣昌, 王亚楠, 赵建夫. 氨氮负荷对膜曝气生物膜反应器部分亚硝化性能的影响[J]. 环境科学学报, 2015,35(4):991-998.
[6] 朱文君, 杨扬, 乔永民, 等. 水力停留时间对自然曝气生物滤床净化效能的影响[J]. 生态科学, 2013,32(2):224-229.
[7] 李民龙, 梁子彦, 汪奇欣, 等. 不同填充率短程硝化泥膜MBBR微生物特征分析[J]. 环境科学与技术, 2022,45(10):106-115.
[8] 陈小红, 许贻斌, 林永青, 等. 不同填料生物膜对海水养殖尾水的脱氮效能及微生物群落分析[J]. 大连海洋大学学报, 2024,39(1):9-19.
[9] Sudarno U, Winter J, Gallert C. Effect of varying salinity, temperature, ammonia and nitrous acid concentrations on nitrification of saline wastewater in fixed-bed reactors[J]. Bioresource Technology, 2011,102(10):5665-5673.
[10] 季必霄. 盐度的快速提升对生物膜硝化能力的影响[D]. 邯郸: 河北工程大学, 2021.
[11] 王玉, 姚倩, 彭党聪, 等. 不同C/N值下SBBR中生物膜硝化特性研究[J]. 中国给水排水, 2016,32(5):23-27.
[12] 郑桂林, 张朝升, 曹勇锋, 等. DO在SBBR生物膜中传递及对同步硝化反硝化的影响[J]. 中国给水排水, 2016,32(3):22-26.
[13] 李韧, 于莉芳, 张兴秀, 等. 硝化生物膜系统对低温的适应特性: MBBR和IFAS[J]. 环境科学, 2020,41(8):3691-3698.
[14] 国家环境保护总局. 水和废水监测分析方法(第四版)[M]. 北京: 中国环境科学出版社, 2002.
[15] 李宁. 低有机负荷条件下EPS在污泥颗粒化进程中的变化规律及其作用机制[D]. 西安: 西安建筑科技大学, 2019.
[16] Felz S, Vermeulen P, Loosdrecht M C V, et al. Chemical characterization methods for the analysis of structural extracellular polymeric substances (EPS)[J]. Water Research, 2019,157:201-208.
[17] Schmidt T, McCabe B K, Harris P W, et al. Effect of trace element addition and increasing organic loading rates on the anaerobic digestion of cattle slaughterhouse wastewater[J]. Bioresource Technology, 2018,264:51-57.
[18] Zheng Z, Li W, Huang X, et al. Effect of trace elements and optimization of their composition for the nitrification of a heterotrophic nitrifying bacterium, Acinetobacter harbinensis HITLi7T, at low temperature[J]. Annals of Microbiology, 2017,67:715-725.
[19] Wagner F B, Nielsen P B, Boe-Hansen R, et al. Copper deficiency can limit nitrification in biological rapid sand filters for drinking water production[J]. Water Research, 2016,95:280-288.
[20] Li C, Liu S, Ma T, et al. Simultaneous nitrification, denitrification and phosphorus removal in a sequencing batch reactor (SBR) under low temperature[J]. Chemosphere, 2019,229:132-141.
[21] 杨思敏, 齐嵘, 杨敏. 低温对生物接触氧化反应器硝化性能的影响[J]. 环境工程学报, 2018,12(11):3028-3033.
[22] 李辰, 何文杰, 黄廷林. 一体式膜生物反应器同步硝化反硝化性能研究[J]. 中国给水排水, 2010,26(5):131-133,137.
[23] 刘思强, 信欣, 杨雯钰, 等. 不同热解温度BC对BAF处理水产养殖废水的影响机制[J]. 中国环境科学, 2023,43(3):1131-1141.
[24] 王萱, 郑俊, 张德伟. 高滤速、低滤层下的反硝化深度脱氮研究[J]. 水处理技术, 2023,49(1):103-107.
[25] Zhao B H, Yang H S, Li Y Q. Achieving partial nitrification in a continuous-flow bioreactor by free ammonia treatment combined with low dissolved oxygen[J]. Biochemical Engineering Journal, 2024,203:109194.
[26] Chang M, Liang B, Zhang K, et al. Simultaneous shortcut nitrification and denitrification in a hybrid membrane aerated biofilms reactor (H-MBfR) for nitrogen removal from low COD/N wastewater[J]. Water Research, 2021,211:118027.
[27] Kasia C, Myszka K. Biosynthesis of extracellular polymeric substances (EPS) and its role in microbial biofilm formation[J]. Polish Journal of Environmental Studies, 2007,16(6):799-806.
[28] Clements E, Nahum Y, Pérez-Calleja P, et al. Effects of temperature on nitrifying membrane-aerated biofilms: An experimental and modeling study[J]. Water Research, 2024,253:121272.
[29] 温沁雪, 施汉昌, 陈志强. 生物膜微环境和传质现象研究进展[J]. 环境污染治理技术与设备, 2006,6:1-5.
[30] 李曼, 祁丽, 彭静波. 纳米塑料对好氧/缺氧生物反硝化脱氮系统的影响探究[J]. 水处理技术, 2024,50(8):134-139,144.
[31] 孙龙, 陈国芬, 雷相杰, 等. 3种滤料组合下生物滞留池的性能和微生物群落的探究[J]. 环境科学与技术, 2024,47(8):61-75.
[32] 杨文焕, 邓子威, 徐岩, 等. 光合细菌对活性污泥微生物群落结构及功能的影响[J]. 中国环境科学, 2024,44(3):1314-1323.
[33] Li Y Y, Liu L, Wang H J. Mixotrophic denitrification for enhancing nitrogen removal of municipal tailwater: Contribution of heterotrophic/sulfur autotrophic denitrification and bacterial community[J]. Journal of Cleaner Production, 2022,814:151940.
[34] 孙峰, 余昕洁, 武威, 等. 基于SNADPR作用的复合式人工快速渗滤系统的运行性能及微生物学特征[J]. 环境工程学报, 2021,15(10):3387-3399.
[35] 董子威, 汪如, 刘莉屏, 等. 微生物菌剂联合填料对污水水体的脱氮效果及其水体中微生物群落结构的影响[J]. 环境工程, 2023,41(S1):596-601.
[36] Yang B, Sun J W, Wang Z Y, et al. Sustainable biochar application in anammox process: Unveiling novel pathways for enhanced nitrogen removal and efficient start-up at low temperature[J]. Bioresource Technology, 2024,402:130773.
[37] 罗镕, 李军, 高鹏, 等. 冬季自然温度下外加AHLs对ANAMMOX生物膜的影响[J]. 中国环境科学, 2024,44(8):4292-4302.
