
 >
>蓝田生物群:一个认识多细胞生物起源和早期演化的新窗口
时间:
摘要蓝田生物群位于安徽省休宁县蓝田镇,保存在埃迪卡拉纪早期蓝田组的黑色页岩中,是已知最古老的复杂宏体生物群,既包含了扇状、丛状生长的海藻,也有具触手和类似肠道特征、形态可与现代腔肠动物相比较的后生动物.这一特殊埋藏的生物群为多细胞生物的起源和早期演化带来了新的启示:微体真核生物在新元古代大冰期结束后迅速演化出宏体形态,它们底栖固着生活在较深水的安静环境中.也许,早期多细胞生物形态复杂化和生物多样性的产生与有性繁殖方式和世代交替现象的出现紧密相关.该时期海洋水体复杂多变的氧化-还原条件,很可能是蓝田生物群繁盛和特殊埋藏的重要原因.
关键词蓝田生物群埃迪卡拉纪多细胞生物起源与早期演化
多细胞生物的出现是地球生命进化史上极为重要的革新事件.生物多细胞化以后,才有细胞的分化,进一步实现器官的分化以及各种功能和形态的出现.在现今生物圈中,包括人类在内的所有肉眼可见的生命,几乎都是多细胞宏体生物,它们在生物谱系树上属于真核生物一枝,也是我们常说的“高等生命”.在地质历史中,自寒武纪至现今,这些高等生命在地球生物圈中发挥了重要的作用,但它们是何时、何种环境背景下以何种形态由单细胞生物演化而来?要回答这些问题,也许只有保存在古老岩层中的生物化石才能提供最直接的证据.
在地球生命史中,多细胞生物有着一段扑朔迷离的早期演化历史.在寒武纪早期,以小壳动物群和澄江动物群为典型代表的化石生物群,显示了多细胞动物在距今5.4至5.2亿年间发生了大规模的辐射[1~3].除一些类型外,大部分现生动物门类在这个时段都有代表性分子出现,这就是通常所说的“寒武纪大爆发”.而在寒武纪之前,虽然多细胞宏体生物化石相对稀少,但一类形态特异的、大型的和软躯体印模保存的埃迪卡拉宏体生物化石在晚前寒武纪(距今5.8~5.4亿年间)的地层中有着广泛的分布[4~9].尽管目前对这一化石组合的生物属性以及与寒武纪以来的多细胞宏体生物的亲缘关系还存在很多争议[10~15],但它们显然都属于多细胞宏体生物,部分类型也可以解释为体型奇特的腔肠动物和软体动物[4,16,17].
迄今为止,最古老的埃迪卡拉生物组合来自加拿大距今5.79~5.65亿年的深水沉积岩石中,称为“阿瓦隆生物群”(AvalonBiota)[8],表明了这个时期的大气圈和海洋中已经含有足够的氧气,从而使得深海区域都能够适合宏体真核生物的生存[18].而在此之前的地球历史中,可靠的宏体真核生物化石极为稀少,大家也普遍认为海水中溶解的氧气不足以支持宏体真核生物的大量发展.
20世纪80年代以来,在中国南方扬子地台早于5.8亿年的陡山沱组地层中,发现了以“瓮安生物群”为代表的磷酸盐化和硅化的微体真核生物化石库,如,在贵州瓮安地区、湖北三峡地区、湖北保康磷矿、江西上饶磷矿等地,不但发现了大量的大型带刺疑源类化石,还发现了保存精美的动物胚胎化石和多细胞藻类化石,它们为探索新元古代大冰期之后、埃迪卡拉生物群出现之前的多细胞真核生物早期演化提供了重要的实证材料[19~34].对这些化石库进行的一系列研究表明,在埃迪卡拉纪早期,包括多细胞藻类和后生动物在内的真核生物,已经发生了辐射.同时,相关地层的稳定同位素地球化学、元素地球化学、矿物学和沉积学研究,揭示了埃迪卡拉纪的环境发生了剧烈变化.尽管埃迪卡拉纪早期的海洋斜坡和盆地都是缺氧的,甚至是硫化的环境,但该时期海洋和大气圈发生了多次氧化事件[35~40].这些研究似乎证实了埃迪卡拉宏体生物群出现之前的浅海生态系统中,真核生物不仅是以微体为主,而且海洋还存在氧化-还原梯度不同的分层现象.
本文介绍的距今约6亿年的“蓝田生物群”,产自中国安徽省南部休宁县蓝田镇埃迪卡拉纪早期蓝田组黑色页岩中,它不但在时代上早于埃迪卡拉生物群,而且生物群面貌和保存方式也显著不同.这一独特的宏体化石生物群,为我们重新认识复杂宏体多细胞生物的早期演化和环境背景打开了一个新窗口.
1蓝田生物群
1.1研究历史
“蓝田生物群”位于安徽省南部休宁县蓝田地区,化石以碳质压膜形式保存在页岩层面.该化石生物群最早由邢裕盛等人[41]进行了报道之后,相继有数篇论文对该地区的宏体化石进行了描述和研究[42~44].1992年阎永奎等人[45]描述了12个属18个种的宏体藻类化石,并把该生物组合定名为“蓝田植物群”;1997年,唐烽等人[46]重点讨论了该化石生物群中的一类圆盘、椭球状化石,认为它们是后生植物性分化的宏体化石证据.1994~1999年,Yuan等人[47]对该生物群进行了系统的野外标本采集和详细的研究,把以前描述的50个属种归入了12~15个种一级的分类单元并对其中的一类球形化石进行了深入的埋藏学研究[48].
近年来,袁训来等人对该生物群进行了再次研究,于2011年2月在Nature上发表了题为“埃迪卡拉纪早期具形态分异的宏体真核生物组合”的科研论文.在同期的新闻和评述栏目,国际早期生命研究专家格·拿邦教授(Prof.GuyNarbonne)撰写了题为“WhenLifeGotBig”的评述,他指出,该化石生物群“是地球上迄今最早的宏体生物”、“是研究早期复杂宏体生命的一个新窗口”[49].
根据新的资料[50],该生物组合是迄今最古老的复杂宏体真核生物群,鉴于类似动物化石的新发现,以及产出时代上的新认识,本文建议把这一特殊埋藏的黑色页岩相的宏体化石生物群更名为“蓝田生物群”.
1.2蓝田生物群简介
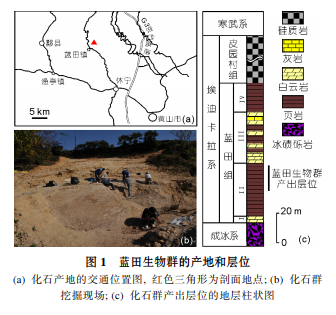
根据化学地层学、层序地层学和事件地层学的研究,结合区域地层对比,蓝田生物群是一个老于所有埃迪卡拉生物群的特殊埋藏宏体真核生物化石群,时代限定在5.8~6.35亿年之间[50],产自新元古代埃迪卡拉系蓝田组的黑色页岩中(图1),化石以碳质压膜的形式保存在页岩的表面,形态保存完整,绝大部分类型具有固着装置,表明它是一个底栖固着生长的复杂宏体生物群.
该生物群包含了扇状、丛状生长的海藻(图2(a)),也有具触手和类似肠道特征、形态可与现代腔肠动物相比较的后生动物(图2(b),(c)).经初步研究,至少能识别出15个不同形态类型的宏体生物(图3).保存化石的页岩微细层理发育,没有发现任何反映强水动力环境的沉积构造和化石搬运迹象,表明这些宏体生物为原地埋藏,生活时应该在最大浪基面之下、透光带之中.根据当时的古地理位置,以及参考现代海洋环境的标准,蓝田生物群大致生活在台地边缘或台地上局部的静水区域,水深在50~200m之间.
从化石的埋藏状态、生物群的总体面貌上看,蓝田生物群都与埃迪卡拉生物群有着明显的区别.蓝田生物群中的化石富含有机质,是以碳质压膜形式保存在页岩的层面上的实体化石,而绝大部分埃迪卡拉生物群中的化石是保存在砂岩或粉砂岩中的软体印模,化石一般不含有机质成分.虽然这两个生物群都是以底栖固着型的宏体生物为主体,但没有发现形态相类似的生物属种.另外,大部分埃迪卡拉生物群(少部分除外,如Avalon生物群)生活在距离海岸较近的浅海区域,而蓝田生物群则生活在静水区域.
已有的众多资料显示,在新元古代发生了一系列重大地质事件,如Rodinia超级大陆的裂解、全球性的大冰期事件等,它们对地球生物圈的演化带来了深刻的影响[51~54].在大冰期之前,浅海生态系统是以原核生物形成的叠层石-微生物席为主体.真核生物虽然在古元古代就已经出现,但演化较为缓慢.化石记录显示,它们主要是个体微小的单细胞藻类(主要是光球疑源类)以及部分形态简单的球状和带状体(主要是chuarid-tawuiid组合)[33].而在新元古代大冰期之后,真核生物得到了快速发展.蓝田生物群就是产自“雪球地球”事件刚刚结束后不久的第一个海侵沉积序列的岩层中.
在地球历史上,大气和海洋中的氧气积累是一个长期和缓慢的过程,自古元古代以来直到新元古代早期,海水明显分层,下层水体为缺氧环境甚至是硫化的海洋[55].新元古代大冰期结束后不久,以真核多细胞生物为主体的“蓝田生物群”就出现在台地边缘或台地深水区域,并且以底栖固着方式、原地生活在黑色页岩沉积相的环境中,说明该时期至少在局部较深水环境中已经含有足够的氧气,能够满足多细胞藻类和后生动物的呼吸作用.另一方面,该生物群的保存没有经过搬运和再沉积作用,以及受其他水动力条件的影响,它们之所以能够以碳质压膜形式完好地产出,推测其原因,很可能与该时期海洋中普遍存在的间歇性缺氧事件有关[50].
埃迪卡拉纪是海洋深水区域由还原状态向彻底氧化过渡的转折时期,也许正是这一独特的环境背景造成了蓝田生物群的生存、死亡和埋藏.该生物群在生活时,底层海水应该都处于富氧状态,而有机碳库的氧化或更深层还原性海水的上涌可能导致了它们的死亡和埋藏.这一氧化和还原背景的转换过程在埃迪卡拉早期甚至到寒武纪很可能循环往复地出现,直到晚古生代海洋才被彻底氧化[56,57].在蓝田组黑色页岩中,化石富集层自下而上重复多次出现,并延续了约15m厚的岩层,显示该生物群经历了多次繁盛、被埋藏和再度繁盛的过程.而且该段黑色页岩岩性均一,没有发生明显沉积相的变化,化石生物几乎都是底栖固着类型,它们生活时需要相对稳定的底质和水体条件.因此,底层海水化学性质(特别是氧含量)的变化很可能是“蓝田生物群”繁盛、被埋藏和再度繁盛的主要原因.
2多细胞生物的起源与早期演化模式
2.1以往的认识
真核生物的起源是早期生命演化史上的一个革新事件,它们与大气圈中自由氧的出现紧密相关.可靠的单细胞真核生物化石可以追溯到古元古代[58],它们的主要枝系在新元古代冰期之前都已经出现[59],例如,杂色藻类以及绿藻、红藻在距今13~10亿年间已经发生了分异[60,61],原生动物在7.5亿年前也有代表分子出现[62].虽然后生动物在新元古代冰期之前可靠的化石记录很少[63,64],但从最保守的分子钟的推算,原口动物和后口动物至少在7亿年前就发生了分异[65],有的分子钟推算甚至估计原口动物和后口动物在10亿年前就已经起源[66~69].早在19世纪,Haeckel就推测过,早期后生动物应该是微体的,形态与现生动物的胚胎与幼虫相类似[70].现代发育生物学家也认为后生动物在获得宏体形态之前,它们类似于现代无脊椎动物幼虫,并且存在较长时间的早期演化历史[71,72].
相对于微体生物而言,多细胞宏体生物对环境条件的要求更高,它们不但需要较为稳定的生存空间,而且介质中氧含量必须达到一定的浓度.宏体生物在埃迪卡拉纪中晚期浅海中广泛出现,并具有较大的形态分异,在相关的地层中还发现了大量的遗迹化石[73~76],另外,在一些深水沉积岩中,也有部分埃迪卡拉生物群被发现,如Avalon生物群[8]、庙河生物群[77],表明该时期的宏体生物(包括后生动物和多细胞藻类)在底栖生态系统中已经扮演了重要的角色.尽管对埃迪卡拉生物群中某些类型的生物属性还存在不同的看法,但它们的广泛分布意味着在埃迪卡拉纪中晚期,以宏体生物为主体的复杂生态系统业已形成,浅海海域已经完全被氧化,而且深水区域也是有氧环境.
相关知识推荐:生物方面的核心期刊
从生物进化的角度来看,多细胞动物在埃迪卡拉生物群之前应该有一个较长的演化历史.但由于化石的稀缺,在埃迪卡拉纪早期及之前的地层中不但没有发现宏体动物化石,也没有发现可靠的遗迹化石,甚至在沉积岩层中没有任何动物搅动的痕迹,因此,大家普遍推测,埃迪卡拉生物群之前的动物应该是微体的、营浮游生活的[78],它们保存为化石的可能性很小.
20世纪末,中国贵州“瓮安生物群”的发现为早期多细胞生物的研究带来了新资料.该化石生物群中距今约6亿年[79],老于埃迪卡拉生物群.早期的磷酸盐化和硅化作用使生物的有机体得以完好地保存,在该生物群中,发现的多细胞生物化石都是微体的,其中包括微体的多细胞藻类、动物胚胎化石以及微体管状腔肠动物化石[19~22,33,80,81].同时在湖北三峡地区的埃迪卡拉系陡山沱组下部的硅质岩中也发现了大量的动物胚胎化石[82,83].部分学者还认为,来自澳大利亚、东欧地台和西伯利亚等地的埃迪卡拉纪早期的某些大型带刺疑源类也有可能属于动物胚胎的休眠期囊胞[84].长期以来,在保存动物胚胎化石的相应层位中并未找到可靠的动物成体,一些学者对这些动物胚胎化石也存在不同的认识[31,85],如解释为团藻、巨大硫细菌或原生生物[31,86~88],但进一步的研究显示,迄今为止,对这些内部具有细胞分裂的球形化石最合理的解释还是动物胚胎化石的观点[29,32,83,89].上述一系列的研究进一步证实了埃迪卡拉生物群出现之前,后生动物应该有较为广泛的分布,也似乎更加让人们确信:该时期的后生动物应该是微体的,并且营浮游生活.
2.2蓝田生物群赋予的新认识
蓝田生物群的新资料为我们重新认识多细胞生物的早期演化打开了一个新窗口.首先,它告诉人们,在埃迪卡拉生物群出现之前已经存在宏体多细胞真核生物,有多细胞藻类,也有后生动物.第二,与埃迪卡拉生物群的生活方式类似,它们绝大多数也是底栖固着型的生物.第三,蓝田生物群中的大部分类型与埃迪卡拉生物群的形态有着明显的差异,也许指示着这两个生物群没有内在的先后演化关系.蓝田生物群中的很多类型可以与其后的多细胞生物对比,其中,分枝的和扇状的多细胞藻类与庙河生物群以及现代的藻类也许存在直接的演化关系;而且那些可能动物化石具有的锥状体型、触手和肠道特征与现生的某些无脊椎动物(如腔肠动物、蠕虫动物等)的体型结构可以比较.
一般认为,早期后生生物应该生存乃至起源于浅水富氧的环境,然而蓝田生物群却生活在较深水并且间歇性缺氧的静水环境,这一独特的现象也许与后生生物的繁殖机制有关.我们知道,后生生物的主要繁殖方式是有性繁殖,雌雄配子的结合如果发生在体外,就需要一个相对比较稳定的水体环境,它们的子代不用进行迁移就能够生活在母体周围较为固定的场所.即使早期后生生物的有性繁殖机制还比较原始,蓝田地区埃迪卡拉纪早期的海洋静水环境也许更加适合这一过程的发生.蓝田生物群中的一些居群个体大小非常类似(图4),它们绝大部分都完好地保存了固着器,表明它们属于原地埋藏的底栖固着类型.对图4中扇状化石进行了统计,它们的长度主要分布在1.5~3.5cm之间,从固着部分向上的发散角主要分布在12°~32°之间,具有明显的正态分布特征(图5(a),(b)),显示它们属于一个种.根据现代遗传学和居群生态学知识,这些形态、大小都类似的同一种生物,并且集中生活在同一环境中,很可能表明它们是有性繁殖产生的同一代居群.这一现象与埃迪卡拉化石Funisiadorothea的繁殖机制非常类似[90].图5对图4中扇状化石的测量(a)为化石个体长度测量的直方图,显示它们主要集中在1.5~3.5cm的范围内;(b)为化石个体发散角测量的直方图,显示它们主要集中在12°~32°
根据蓝田生物群的总体特征以及相关的环境信息,对多细胞生物的起源、早期演化及其环境背景也许可以做如下的推测:
新元古代大冰期之前的浅海底栖生态系统是以原核生物为主体,真核生物虽然在古元古代就已经起源,但由于氧气含量较低带来的一系列环境因素的影响,延缓了真核生物的多样化进程,它们分异度较低,以微体类型为主,大部分营浮游生活在水体的浅表层含氧带.经过长达一亿多年的新元古代冰期间冰期事件之后,大气圈氧含量明显增加,海洋的深层水被逐渐地氧化,一些浮游的微体真核生物能够迁移到较深水的海底生活,并建立了以多细胞生物为主体的高级生态系统.与多细胞藻类一样,这个时期的后生动物也是营底栖固着生活,它们类似于现代的腔肠动物或海绵动物,没有对沉积物产生任何搅动作用.在最大浪基面之下的有光带静水环境中,这些藻类和动物有可能都是进行有性繁殖,从而大大提高了遗传物质的变异,并进一步导致了形态的复杂化和多样化.在这样的环境中,多细胞生物,特别是动物经过了数千万年的演化,它们的体型结构以及繁殖机制逐渐完善.在埃迪卡拉纪中晚期,它们逐步迁移和扩散到较浅水的近岸环境中.——论文作者:袁训来①,陈哲①,肖书海①②,万斌①,关成国①,王伟①,周传明①,华洪③
