
 >
>黄河三角洲盐碱地不同植被模式的土壤改良效应
时间:
摘 要 黄河三角洲是我国滨海盐碱地的重要分布区,种植植被是盐碱地绿色改良的主要生态修复措施。 为探讨滨海盐碱地不同植被模式的土壤改良效应,探索适宜植被模式,选取黄河三角洲盐碱地竹柳+NyPa 草、旱柳+NyPa 草、柽柳+紫花苜蓿、白蜡+柽柳+紫花苜蓿 4 种林草措施为研究对象,以纯竹柳为对照,测定土壤水分物理参数、盐碱含量、土壤养分及微生物数量等 20 个指标,并利用主成分分析、聚类分析和模糊数学隶属函数等统计方法评价了不同植被模式的土壤改良效应。 结果表明: 林草复合模式可显著改善滨海盐碱地的土壤理化性能,增加土壤孔隙度和贮水量,降低土壤密度,提高土壤有机质、速效养分含量和土壤微生物数量。 其中,白蜡+柽柳+紫花苜蓿的乔灌草混交模式在压碱抑盐、增加土壤养分和微生物数量的效果最好,而旱柳+NyPa 草的乔草混交模式改良土壤水分物理性能的效果最好。 不同植被模式对黄河三角洲滨海盐碱地的综合改良效应表现为白蜡+柽柳+紫花苜蓿>旱柳+NyPa 草>竹柳+NyPa 草>柽柳+紫花苜蓿。
关键词 林草措施; 土壤孔隙; 土壤养分; 土壤微生物; 土壤盐分
土壤盐渍化是限制我国土地生产力提高的主要因子之一,我国盐渍土总面积约为 3.6×107hm2,占全国可利用土地面积的 4.9%[1]。 滨海盐碱地是我国盐碱土的主要组成部分,2×104km2的沿海滩涂和1.8×104km 的滨海地带与岛屿沿岸广泛分布着各种滨海盐土,总面积可达 5×106hm2 [2]。 滨海盐碱地是一种重要的土地资源,在扩大耕地资源和改善泥质海岸带生态环境方面潜力巨大。 黄河三角洲是由黄河携带大量泥沙在入海口淤积而成的新生陆地,受河流和海洋等多种动力交互作用,该地区潜水埋深浅、矿化度高,导致土壤盐渍化较重、范围广[3-4]。盐碱地土壤改良及合理利用是黄河三角洲高效生态经济区建设亟需解决的重点问题,也是扩大黄河三角洲土地资源利用的关键环节。
土壤物理性状、盐碱状况、养分含量及微生物数量等是衡量土壤质量健康与否的主要指标[5-7]。 土壤物理性状对土壤的水、肥、气、热及其化学和生物学过程起着重要的调控作用[5,8],土壤养分状况对植物的生长发育和分布有直接影响[4,9-10]。 土壤微生物参与地球化学元素循环,其种群数量和分布是反映土壤生物稳定性的一个显著指标,对植物养分有效利用和土壤结构的形成与改良等有重要影响[5,7,11-12]。 植物与土壤之间存在着复杂的交互效应,土壤物理性质及养分状况的变化直接影响植物生长和土地生产力,不同植被类型对盐碱地土壤质量的影响差异较大[4-5,13]。 盐碱地土壤养分、水分的有效性受到土壤盐分的显著影响,导致不同植被类型间土壤酶、 微生物与养分之间关系较为复杂[6,10,12]。 然而,在盐碱生境下,不同林草复合模式对土壤微生物活动、土壤有效养分形成及提升地力等方面的作用过程与机制尚不清晰,这在一定程度上阻碍了植被复合系统在滨海盐碱地土壤改良上的应用。
黄河三角洲土壤盐渍化严重、土壤贫瘠且通气透水性差,严重限制了植被的生长发育和分布格局[4]。 目前针对黄河三角洲盐碱地改良与利用问题,主要采取了暗管排盐等水利工程措施,掺沙、覆盖等物理措施,添加牛粪、石膏、秸秆等化学改良措施,以及种植耐盐碱植物等生物改良措施[14-15],其中筛选和培育耐盐碱植物及其栽植模式是一种改良盐碱地的重要技术。 植被措施改良盐碱地具有可持续强和地力能自我维持等优点,而植物材料和植被模式选择是生物改良措施的关键环节。 林草复合系统构建作为一种重要的退化生态系统的修复重建技术,在植被生态恢复和实现可持续发展方面发挥的生态、经济效益潜力巨大[16-17]。 我国林草复合系统构建研究主要集中在西北、 华南和西南三大区域[18],其在提高土地生产力、改善生态环境和高效利用自然资源等方面具有明显优势,可较好促进林业⁃草业系统的相互融合及对退化生境的综合治理。目前,国内外许多学者已开展了单一树木、草本种植以及造林模式改良盐碱地方面的研究[6,13],主要以探讨土壤盐碱指标变化为主,但涉及植被复合模式对滨海盐碱地土壤改良效应的研究较少,在一定程度上制约了盐碱生境下植被复合系统的模式筛选和结构优化。
鉴于此,本研究在对重度盐碱地进行传统工程措施整地的基础上,以耐盐碱牧草形成的林草混交和纯竹柳为研究对象,测定分析不同植被模式下的土壤水分物理参数、盐碱含量、速效养分以及微生物数量等,结合主成分分析、聚类分析和模糊数学综合评价法,探讨不同植被模式对滨海盐碱土的改良效应,明确滨海盐碱地改良的适宜植被模式,以期为黄河三角洲盐碱地绿化植被模式构建提供理论依据和技术参考。
1 研究区域与研究方法
1 1 试验地概况
研究地点位于山东省东营市河口区的黄河三角洲综合训练基地内(37°53′51″ N, 118°48′22″ E),试验区域面积为 2.5 hm2。 该区属于暖温带半湿润地区,大陆性季风气候,年均气温 12.1 ℃ ,无霜期长达201 d,≥10 ℃ 的积温约 4200 ℃ ,年降水量 500 ~600 mm,多集中在夏季,7—8 月降水量约占全年降水量的一半,年蒸发量 1700 ~ 1800 mm。 试验区地下水位浅、矿化度高、地势平坦,滩涂土壤为冲积性黄土母质,机械组成以粉砂和淤泥质粉砂为主,沙粘相间,易于压实,通气透水性差。 人工林主要以刺槐(Robinia pseudoacacia)、白蜡(Fraxinus chinensis)、竹柳(Salix americana)、旱柳( Salix matsudana)、柽柳(Tamarix chinensis)等树种为主,草本植被主要以芦苇(Phragmites australis)、翅碱蓬( Suaeda salsa)、白茅(Imperata cylindrica)、獐毛(Aeluropus littoralis)和菌陈蒿(Artemisia capillaris)等耐盐植物为主。
1 2 研究方法
1 2 1 样地设置与样品采集 2014 年 4 月在试验地内进行工程整地,修筑高 2.0 m、宽 50 m 的台田,平整土地淡水压盐碱后 0 ~ 40 cm 土层的土壤含盐量为 0.25%,pH 为 8.3。 试验构建以下 4 种林草混交模式:竹柳+NyPa 草(Distichlis spicata) (ZN),旱柳+NyPa 草 ( HN), 简称乔 草 模 式; 柽 柳 + 紫 花 苜 蓿(Medicago sativa)(CZ),简称灌草模式;白蜡+柽柳+紫花苜蓿(BCZ)简称乔灌草模式。 以纯竹柳为对照(CK)。 除柽柳+紫花苜蓿的株行距为 1.5 m×2.0 m之外,其他 4 种植被模式的株行距均为 2. 0 m × 3.0 m。 每种植被模式带长 100 m、宽 10 m。 试验采用随机区组设计,每种林草模式 3 次重复。 试验的2 种牧草分别为紫花苜蓿和 NyPa 草,其中紫花苜蓿种植采用条播方式,播种深度为 1.5 cm,行距30 cm,播种量为 20 kg·hm-2;NyPa 草采用根蘖分 7~10 株进行 栽 植, 栽 植 深 度 为 10 cm 左 右, 株 行 距 为20 cm×40 cm。 NyPa 草和紫花苜蓿在种植第 1 年任其自然生长,以后每年在 6 月底、8 月初分 2 次进行刈割。
2017 年10 月对不同植被模式下的树木生长状况进行调查(表 1),同时采集土壤样品,在各植被模式的试验样地内,按 S 型布设 5 个试验样点,每样点采集根系集中分布层 0~40 cm 深度的土壤样品 2 份,一份风干后用于测定土壤理化性质;另一份装于密闭自封袋,4 ℃冰箱保存,用于测定土壤微生物数量。
1 2 2 测定方法 采用烘干法测定土壤含水量,环刀浸水法测定土壤密度、孔隙度、贮水性等各项水分物理指标,土壤孔隙比为孔隙体积与土粒体积的比值;pH 采用 pH 计(水土比 5 ∶ 1),可溶性盐采用重量法测定(水土比 5 ∶ 1);土壤有机质采用重铬酸钾容量法,速效氮用碱解⁃扩散法,速效钾用 NH4Ac 浸提⁃火焰光度法,速效磷采用 NaHCO3浸提⁃钼锑抗比色法[19]。土壤微生物区系的分析采用稀释平板法,细菌采用牛肉膏蛋白胨培养基,真菌采用马铃薯葡萄糖琼脂培养基,放线菌采用改良高氏 1 号培养基[20]。
1 3 数据处理
采用 Excel 2000 和 SPSS 18.0 软件对数据进行统计分析,数据符合正态分布。 土壤理化指标采用单因素方差分析(one⁃way ANOVA)检验(α = 0.05),相关性采用 Pearson 相关分析,并进行主成分分析和聚类分析。 图表中数据为平均值±标准差。
2 结果与分析
2 1 土壤水分物理特征
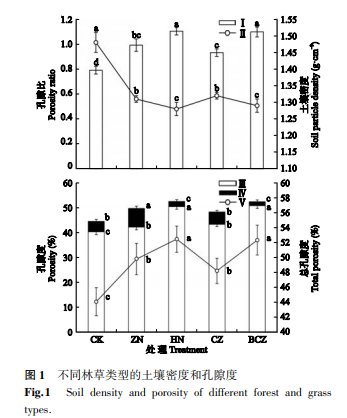
2 1 1 土壤密度和孔隙度 由图 1 可知,不同林草混交模式下的土壤密度介于 1.28 ~ 1.32 g·cm-3,土壤孔隙比介于 0.93 ~ 1.10,而纯竹柳土壤密度最高达 1.48 g·cm-3,孔隙比最低为 0.79。 林草混交土壤密度显著低于纯竹柳样地土壤,与竹柳相比,竹柳+NyPa 草、旱柳+NyPa 草、柽柳+紫花苜蓿、白蜡+柽柳+紫花苜蓿的土壤密度分别降低 11.5%、13.5%、10.8%、12.8%。 土壤孔隙比、毛管孔隙度、总孔隙度大小均表现为白蜡+柽柳+紫花苜蓿、旱柳+NyPa草较高,其次是竹柳+NyPa 草、柽柳+紫花苜蓿,显著高于竹柳。 不同植被模式下土壤总孔隙度为 44.1% ~52.5%,毛管孔隙度为 40.3% ~50.8%,非毛管孔隙度为 1.5% ~7.5%。 与竹柳相比,竹柳+NyPa 草、旱柳+NyPa 草、柽柳+紫花苜蓿、白蜡+柽柳+紫花苜蓿的总孔隙度分别增加 12.9%、18.9%、9.3%、18.6%。 林草模式可显著改善盐碱地的土壤孔隙状况,总体表现为土壤孔隙度增大,土壤密度减小。 但不同植被模式的土壤孔隙状况改善情况差异较大,林草模式土壤孔隙的改善状况显著好于纯竹柳,其中白蜡+柽柳+紫花苜蓿的乔灌草混交和旱柳+NyPa 草的乔草混交模式效果较好。
2 1 2 土壤贮蓄水分特征 由表 2 可知,与竹柳相比,白蜡+柽柳+紫花苜蓿、旱柳+NyPa 草、柽柳+紫花苜蓿、竹柳+NyPa 草 4 种植被模式的土壤含水量分别增加 47.8%、36.8%、31.6%、21.7%。 不同植被模式的土壤贮水量差异显著,其中总贮水量表现为旱柳+NyPa 草和白蜡+柽柳+紫花苜蓿最高,分别比纯竹柳提高 18.9%和 18.6%,土壤贮蓄和调节水分能力最强;其次是竹柳+NyPa 草和柽柳+紫花苜蓿,分别比纯竹柳提高 12.9%和 9.3%。 毛管贮水量也表现为白蜡+柽柳+紫花苜蓿和旱柳+NyPa 草最高,且二者差异不显著,表明该乔灌草混交模式和乔草模式具有贮存供植物生长所需水分的潜能;而竹柳的毛管贮存水分能力较弱。 竹柳+NyPa 草的非毛管贮水量最高,涵养水源潜在功能较强,而乔灌草模式滞留贮存自由重力水的能力较弱。 不同植被模式的涵蓄降雨量和有效涵蓄量差异显著,其中涵蓄降雨量比纯竹柳提高 5.4% ~13.5%。 与竹柳相比,旱柳+NyPa 草、白蜡+柽柳+紫花苜蓿、柽柳+紫花苜蓿、竹柳+NyPa 草 4 种植被模式的有效涵蓄量分别增加22.6%、21.7%、2.7%、1.2%;对降雨的土壤调蓄能力表现为旱柳+NyPa 草、白蜡+柽柳+紫花苜蓿最强,柽柳+紫花苜蓿、竹柳+NyPa 草较弱。
2 2 土壤盐碱含量和养分特征
2 2 1 土壤 pH 和含盐量 由图 2 可知,不同植被模式的土壤含盐量和 pH 差异显著,其中 pH 为 7.6 ~8.4,土壤含盐量为 0.1% ~ 0.3%。 pH 大小由低到高依次为白蜡+柽柳+紫花苜蓿、柽柳+紫花苜蓿、旱柳+NyPa 草、竹柳+NyPa 草、竹柳;土壤含盐量由低到高依次为白蜡+柽柳+紫花苜蓿、旱柳+NyPa 草、柽柳+紫花苜蓿、竹柳+NyPa 草、竹柳。 土壤脱盐率表现为乔灌草模式最高,可高达 65.6%;其次为旱柳+NyPa 草、柽柳+紫花苜蓿、竹柳+NyPa草,分别为43.8%、34.4%、29.7%。 乔木、灌木与牧草的复合混交模式具有一定的压碱抑盐效应,但不同植被模式间差异较大,总体表现为林草混交模式好于纯竹柳,其中乔灌草混交模式压碱抑盐的效果显著好于柽柳+紫花苜蓿和旱柳+NyPa 草,而竹柳+NyPa 草的乔草混交模式降盐抑碱效果较差。
2 2 2 土壤有机质和速效养分含量 由图 3 可知,不同植被模式可显著提高盐碱地的土壤有机质,其中白蜡+柽柳+紫花苜蓿、旱柳+NyPa 草、柽柳+紫花苜蓿、竹柳+NyPa 草的土壤有机质含量分别是纯竹柳的 2.7、2.5、2.0、1.7 倍。 白蜡+柽柳+紫花苜蓿和旱柳+NyPa 草的土壤有机质含量最高,其次是柽柳+紫花苜蓿和竹柳+NyPa 草。 不同植被模式的土壤N、P、K 速效养分含量差异均显著,土壤速效 K 含量最高,其次是速效 P,而速效 N 含量较低。 白蜡+柽柳+紫花苜蓿、旱柳+NyPa 草、柽柳+紫花苜蓿和竹柳+NyPa 草的土壤速效 N 含量是纯竹柳的 2.2、2.0、1.8、1.2 倍;速效 P 含量表现为白蜡+柽柳+紫花苜蓿>竹柳+NyPa 草>柽柳+紫花苜蓿>旱柳+NyPa 草>竹 柳;白蜡+柽柳+紫花苜蓿、旱柳+NyPa草、柽柳+紫花苜蓿、竹柳+NyPa 草的速效 K 含量分别是纯竹柳的 1.9、1.9、1.5、1.4 倍。 不同植被模式可显著提高土壤有机质和速效养分含量,但不同植被模式间表现出较大差异,其中乔灌草混交模式提升土壤肥力效果最好,其次是乔草、灌草混交模式。
2 3 土壤微生物数量
由表 3 可知,不同植被模式下土壤细菌、真菌和放线菌数量差异显著,其中细菌数量在微生物总数中含量最高,占微生物总数比例达 87.3% ~ 92.9%;其次是适宜微碱性环境的放线菌,占微生物总数比例达 3.7% ~ 7.1%;而适宜酸性环境的真菌数量最低,占微生物总数比例仅为 1.5% ~ 4.9%。 不同植被模式显著增强了盐碱地土壤的微生物数量,细菌数量在乔灌草混交模式中最高,其次是竹柳+NyPa 草和柽柳+紫花苜蓿 2 种林草混交模式,旱柳+NyPa草次之,而纯竹柳模式含量较低。 真菌数量表现为旱柳+NyPa 草模式最高,白蜡+柽柳+紫花苜蓿次之,而竹柳+NyPa 草、柽柳+紫花苜蓿和竹柳模式较低。 放线菌数量表现为白蜡+柽柳+紫花苜蓿>旱柳+NyPa 草>柽柳+紫花苜蓿>竹柳+NyPa 草>竹柳。 林草混交可显著提高土壤微生物数量,微生物总量白蜡+柽柳+紫花苜蓿、竹柳+NyPa 草、柽柳+紫花苜蓿、旱柳+NyPa 草 4 种模式分别是纯竹柳模式的2.7、2.4、2.4、2.1 倍。 盐碱生境下的林草混交模式对提高细菌数量较为有利,而对真菌的影响效果较差。
2 4 土壤改良效应的主成分分析和聚类分析
不同植被模式的土壤理化性质差异较大,并且土壤理化参数之间存在着交互作用。 对这些指标进行主成分分析(表 4),可归纳出不同植被模式改良土壤效应的主要指示因子。 由表 4 可知,前 2 个主成分的累计贡献率为 92.5%,能够反映测试指标的大部分信息,因此取前 2 个符合综合数值分析要求的主成分。 第 1 主成分的贡献率为 75.4%,是最大主分量,因子负荷量较大的为土壤有机质、速效K、含水量、含盐量、孔隙比、毛管孔隙度、总贮水量和毛管贮水量,该类可表述为土壤养分、土壤水盐和土壤通气性。 第 2 主成分负荷量较大的为速效 P 和非毛管贮水量。
依据上述 20 项指标对 5 种植被模式进行聚类分析,聚成 3 类,可较好反映该盐碱地植被模式的改良土壤效应(图 4)。 第Ⅰ类包含旱柳+NyPa 草的乔草混交模式和白蜡+柽柳+紫花苜蓿的乔灌草混交模式,其改良土壤水分物理参数及提升肥力效应较强;第Ⅱ类包含竹柳+NyPa 草和柽柳+紫花苜蓿 2种乔木、灌木草本混交模式,第Ⅲ类是纯竹柳,改良土壤效应较弱。
2 5 不同林草模式改良土壤效应的综合评价
依据主成分分析结果,选用因子荷载值较大的10 个指标用来评价不同植被模式的改良土壤效应,分别为土壤有机质、速效 K、含水量、含盐量、孔隙比、毛管孔隙度、总贮水量、毛管贮水量、速效 P 和非毛管贮水量。 由主要因子的隶属函数综合值可知(表 5),不同植被模式改良土壤效应的综合评价为白蜡+柽柳+紫花苜蓿>旱柳+NyPa 草>竹柳+NyPa草>柽柳+紫花苜蓿>竹柳。
3 讨 论
3 1 不同植被模式的蓄水保土功能
林草复合模式具有的多层次植被覆盖可保持水土,特别是草本植物的密集根系可固结土壤,提高土壤表面的抗冲刷能力,能有效减少土壤流失[22-23]。林草复合模式的土壤水分与土壤孔隙结构、植物蒸腾耗水以及蒸降比等密切相关[24]。 滨海盐碱土孔隙度小,土壤易板结,透水、透气性差,改良滨海粘质盐土的关键是增加土壤大孔隙数量和比例,提高土壤饱和导水率[14]。 本研究发现,林草混交可显著改善土壤孔隙结构,土壤密度降低,土壤孔隙度和孔隙比增大,贮蓄水分能力增强。 白蜡+柽柳+紫花苜蓿、旱柳+NyPa 草模式改善土壤孔隙度效果最好,土壤总 贮 水 量 分 别 比 纯 竹 柳 模 式 提 高 18. 9% 和18.6%,孔隙比大于 1.0,属于疏松的高压缩性土,有利于拦蓄水分和减缓径流冲刷,这与牧草根系密集和树木根系具有较强的穿插作用有关[25],同时与牧草、树木枯落物分解形成的腐殖质层也有较大关系[26]。 紫花苜蓿和 NyPa 草根系发达,须根多而稠密,对土壤的穿插和挤压作用较强;柽柳新生枝条和叶片生物量较大,易形成厚的枯落物层,并且柽柳根系扎根深度变化范围较大[27]。 因此,竹柳 +NyPa草、柽柳+紫花苜蓿模式可有效改善土壤孔隙状况,从而促进土壤贮蓄水分的能力,2 种林草模式的土壤总贮水量分别比纯竹柳模式提高 12.9%和 9.3%。黄河三角洲盐碱地道路防护林降低土壤容重可达10% ~ 20%,增加土壤孔隙度可达 4% ~ 10%[28];耐盐碱牧草降低内陆盐碱地土壤容重可达 6% ~ 8%,提高土壤孔隙度可达 5% ~ 6%[25];而本研究林草混交模式与纯竹柳模式相比降低土壤密度 10. 8% ~13.5%,提高土壤总孔隙度 9.3% ~ 18.9%,可见林草混交模式在改善盐碱地土壤通气透水性方面好于单一纯竹柳。 林草复合模式可通过植株腐烂分解提供大量有机质和氮素[23],调节土壤结构类型,从而增强土壤的渗透性和蓄水力。 本研究也发现(表 6),土壤有机质与土壤密度呈显著负相关,与土壤孔隙比、总孔隙度、毛管孔隙度、总贮水量、毛管贮水量呈显著正相关,与土壤含水量、含盐量分别呈极显著正相关和负相关;速效 N 与毛管孔隙度、土壤含水量、毛管贮水量、土壤有机质呈显著正相关,与 pH 和土壤含盐量呈显著负相关;速效 K 与土壤孔隙比、总孔隙度、总贮水量、有效涵蓄量和速效 N 呈显著正相关,与毛管孔隙度、土壤含水量、毛管贮水量、土壤有机质呈极显著正相关,与土壤含盐量呈显著负相关;而速效 P 与相关指标的相关性不显著。 可见,随着土壤有机质和速效养分的提高,滨海盐碱土通气透水性、贮蓄水分和降盐改土能力明显提高,但各指标之间的相关性呈现一定的差异。——论文作者:孙 佳1,2 夏江宝2∗ 苏 丽1,2 赵西梅2 陈印平2 岳喜元2 李传荣1
