
 >
>多肉植物红司(Echeveria nodulosa)花粉离体萌发和花粉管 生长特性研究
时间:
摘 要:以 10%蔗糖+0.5%琼脂为基础培养基,采用花粉离体萌发法研究硼酸浓度、培养温度和培养时间对多肉植物红司(Echeveria nodulosa)花粉萌发和花粉管生长的影响。结果表明:硼酸浓度、培养温度、培养时间三因素以及相互间的交互效应对花粉萌发率的影响差异极显著;硼酸浓度、培养温度以及二者间的交互效应对花粉管伸长的影响显著,而培养时间对花粉管伸长的影响不显著。12~28 ℃有利于提高花粉管萌发率,20~36 ℃有利于花粉管的伸长。三因素交互效应显示:60 mg/L 硼酸+12 ℃+16 h 为红司花粉萌发最佳条件,萌发率为 52.43%。30 mg/L 硼酸+36 ℃+16 h 为花粉管生长适宜条件,花粉管平均长度达 527.74 μm。
关键词:多肉植物;红司;培养温度;花粉管生长
多肉植物红司(Echeveria nodulosa)属于景天科(Crassulaceae)拟石莲属(Echeveria)原生种,原产于墨西哥,叶片上分布着奇特的红色纹路,花粉量丰富,花期 5—7 月。红司能较好适应高低温环境,且观赏性较好,引进国内后进行大量的商品化生产。在育种方面,红司也是一种重要的育种父本材料,曾育出园艺品种 E. shaviana× E. nodulosa 和 E.setosa var. ciliata×E. nodulosa。拟石莲属是景天科中最具有代表性的一个属,其种类比例占到景天科原种的 97%,而分布在墨西哥的拟石莲属多肉植物拥有丰富的多样性,85% 的物种生长在墨西哥[1-2]。此外,拟石莲属多肉植物的形态特征具有较高的吸引力,被世界园艺家和收藏家认为是观赏价值很高的园艺植物。
González-Mancera 等[1]通过电镜扫描拟石莲属花粉,发现大多数品种花粉表现出相似的特征,大小为 27~42 m,外形为球状或椭圆形,表面具有网状或条形纹饰,但有一些表现出不同的特征,如花粉表面具有孔沟或多个萌发孔。 Jimeno-Sevilla 等 [3] 研究发现拟石莲属植物 E. rosea 具有一种特殊的蜂鸟授粉系统,在自然条件下可以获得较高的结果率。但实际上拟石莲属的多肉植物被园艺家引种栽培在室内,缺少有效的传粉者,自花授粉结实率普遍较低,只能采取人工辅助授粉方式对其进行遗传改良,形成具有较高观赏价值的园艺新品种。人工授粉的方式可有效提高育种成功率,Parra-Tabla 等[4]研究发现人工授粉 E. gibbiflora 产生的种子比自然授粉多 1.74 倍。花粉活力是影响人工授粉成功的关键因素之一,而植物的遗传起源是影响花粉活力的重要因素,突变体或种间杂交高度不育[2]。低活力的花粉会影响杂交中可存活种子的数量,只有高活力的花粉在柱头上才有较高的萌发率,能快速伸长到达子房成功完成受精,是人工辅助授粉及杂交授粉的重要保障,因此,研究花粉的活力对育性和结实率具有重要意义[5-6]。在研究花粉活力的过程中,离体萌发法被认为是检测花粉活力快速、可靠且稳定的方法,检测结果与在柱头上的花粉活力相似[7]。本研究为了摸清多肉植物红司花粉活力及花粉管生长特性,以便育种上能更加高效地运用红司培育新品种,采用离体萌发法研究硼酸浓度、培养温度、培养时间三因素及其相互间的交互效应对于红司花粉萌发率及花粉管生长的影响,以期筛选出花粉萌发及花粉管生长的最佳培养条件,揭示红司花粉育性,从而提高育种效果和加速育种过程,并为多肉植物育种方提供依据。
1 材料与方法
1.1 材料
供试材料红司为 2015 年引种种植于贵州省农业科学院园艺研究所多肉种质资源保育棚。取样植株的种植年限为 4 年,选取发育正常、长势健壮、无病虫害且生长情况接近的红司 5 株,并分别编号为 1~5 号,插上标签。取材时间为 2019 年 7 月,晴天早上 9:00—10:00 之间,分别采集 1~5 号红司植株花朵,每株植株采集 5 朵,采集的花粉均为花朵刚开放且柱头出现粘液时期的花粉。
1.2 方法
1.2.1 花粉离体培养 用镊子将花粉轻轻剥落至滴有蒸馏水的离心管中混匀制成悬浮液。以 0.5%琼脂(Agar)+10%蔗糖(Sucrose)为基础培养基,添加不同浓度的硼酸,制成的培养基倒入玻璃培养皿中冷却凝固。分别取 100 µL 的花粉悬浮液滴入固体培养基中,置于恒温培养箱内进行不同处理的暗培养。将培养基置于显微镜进行观察,统计花粉萌发率和测量花粉管长度。空白对照 CK 为 0.5%琼脂( Agar) +10%蔗 糖(Sucrose)为基础培养基,滴入花粉悬浮液后不培养直接观察。
1.2.2 硼酸浓度设置 变量因素 A 为硼酸。分别添加 A1(0 mg/L)、A2(30 mg/L)、A3(60 mg/L)、 A4(100 mg/L)4 种浓度水平的硼酸。
1.2.3 培养温度设置 变量因素 B 为培养温度,设定为 5 种水平,设计覆盖了低中高 3 个温区,分别是 B1(12 ℃)、B2(20 ℃)、B3(28 ℃)、 B4(36 ℃)、B5(44 ℃)。
1.2.4 培养时间设置 变量因素 C 为培养时间,设定为 4 种水平,分别是 C1(1 h)、C2(3 h)、 C3(16 h)、C4(24 h)。
1.2.5 离体萌发率统计和花粉管长度的测量 试验设计为三因素多水平处理,4×5×4 共 80 个组合。在显微镜下随机观察,以花粉管长度大于花粉直径为花粉萌发标准[8]。每个组合重复 3 次,每次的观察视野为 3 个。每个组合观察视野的花粉粒总数为 350~780 粒,统计花粉萌发率(花粉萌发率=萌发的花粉数/花粉粒总数×100%,结果取平均值)。同时,用显微镜微尺测量花粉管长度,每个组合观察视野的花粉管数量为 30~80 个,计算其平均值。
1.3 数据处理
利用 Excel 软件进行数据整理,用 SAS 9.0 软件对组合进行差异显著性、多元相关性及聚类分析。
2 结果与分析
2.1 三因素对红司花粉萌发率的影响
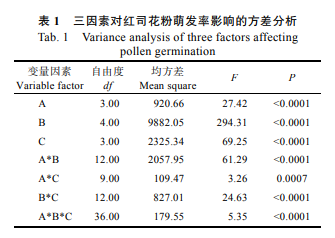
如表 1 所示,硼酸浓度、培养温度、培养时间对红司花粉萌发率均有极显著影响。空白对照 CK 的花粉萌发率仅为 3.71%。从表 2 可见, 60 mg/L 硼酸浓度下花粉萌发率最高,为 15.87%。 12 ℃低温条件下花粉萌发率最高,为 21.86%;花粉萌发率随着培养温度的升高而降低,44 ℃高温下会抑制花粉萌发。培养 16 h,花粉萌发率达到最大值 18.70%,显著高于其他 3 个时间点;培养时间增加至 24 h 时,观察视野中许多花粉管出现破裂,扭曲交缠,不利于统计萌发数量,导致萌发率在数据上出现下降。
从表 3 可见:60 mg/L 硼酸+12 ℃培养 3 h、 16 h,花粉萌发率达到 48.59%、52.43%;两因素的组合中,60 mg/L 硼酸+12 ℃(A3B1)的培养条件下,萌发率为 38.43%,显著高于其他水平组合;硼酸浓度与时间的最佳组合为 60 mg/L 硼酸+16 h(A3C3),萌发率为 20.67%;培养温度与时间的最佳组合为培养 12 ℃+16 h(B1C3),萌发率为 31.43%。硼酸酸浓度、培养温度、培养时间三因素组合条件下花粉的萌发率大于双因素组合。
2.2 三因素对红司花粉管生长的影响
硼酸浓度和培养温度及二者间的交互效应对花粉管生长存在极显著影响(表 4)。空白对照 CK 的红司花粉管平均长度仅为 44.40 μm。培养基中加入不同浓度的硼酸可促进花粉管生长, 30 mg/L 硼酸浓度为最佳处理水平,花粉管平均长度达到 223.22 μm(表 5、图 1)。低温对花粉管的伸长有抑制作用,12 ℃低温培养下花粉管生长过慢,平均长度值仅为 100.2 μm,20~36 ℃为红司花粉管生长适宜温度区间(表 5、图 2);培养时间对花粉管生长的影响并不显著(表 4),培养时间 3~16 h 为花粉管快速生长阶段,培养 16 h 时花粉管平均长度达到 194.33 μm(表 5)。
低浓度硼酸在高温培养条件下对花粉管伸长有促进作用。30 mg/L 硼酸在 36 ℃条件下分别培养 3、16、24 h,花粉管平均长度分别达到了 453.16、527.74、571.52 μm(表 6),但培养 24 h 后 400 μm 以内花粉管数量明显减少(表 7),不利于授粉成功率提高,因此,花粉管生长最佳处理水平为 A2B4C3。
2.3 三因素的交互效应对花粉管生长影响的聚类分析
采用类平均法将 OB1~OB80 共 80 个组合聚为 3 类,阈值为 1.3193。如表 7 所示:第 1 类中,花粉管长度在 400~1100 μm 的花粉比例最高,平均数量为 35.3 个,200~400 μm 的花粉平均数量为 8.0 个,200 μm 以内的花粉平均数量为 6.3 个;第 2 类中,200 μm 以内的花粉比例最高,平均数量为 75.1 个;其次为 200~400 μm,花粉平均数量为 11.7 个,400~1100 μm 的花粉仅为 2.6 个。第 3 类中,花粉管长度主要在 200 μm,平均数量为 30.45 个,其次为 200~400 μm 的花粉,平均数量为 5.7 个,400~1100 μm 的花粉平均数量为 2.9 个。说明 30 mg/L 硼酸在 36 ℃培养 3~16 h 后,对花粉管的伸长有明显的促进作用。
2.4 三因素与红司花粉萌发和花粉管生长的相关性
硼酸浓度、培养温度、培养时间多个变量对花粉萌发和花粉管生长影响的相关性分析发现(表 8):花粉萌发率与培养温度呈极显著负相关(P<0.01),相关系数值为‒0.6044;花粉萌发率、花粉管生长与硼酸浓度和培养时间均呈正相关关系,但相关性均不显著。两组变量的典型相关性分析也得到了相同结果,证实多个变量因素中培养温度(B)与花粉萌发率(y)极显著相关,逐步回归分析获得二者的最优方程 y=31.06-0.58 B (R2 =0.3773),方程极显著(P<0.01)。说明温度是影响红司花粉萌发率最主要的因素,随着温度的增加,花粉萌发率表现为下降。此外,一定范围内硼酸浓度值对萌发和花粉管生长是有利的。
3 讨论
影响花粉离体萌发率的因素除了培养基、温度、硼酸浓度等因素外,还应该考虑取材的一致性,应尽量减少取材的次数,取回的多个花朵制成花粉混合悬浮液,让每个处理都能在材料上保持基本一致。另外,较少的处理组合很难反应花粉萌发及花粉管生长的真实状况,应尽量采用多因素多水平组合处理。0.5%琼脂(Agar)+10%蔗糖(Sucrose)为基础培养基,滴入花粉悬浮液后不培养直接观察,可摸清楚花粉在培养前的萌发状态。本试验中,花粉在空白对照处理下花粉有 3.71% 极少量的萌发,花粉管平均长度仅为 44.40 μm。
硼元素对花粉管生长具有调节作用,在构建花粉细胞壁方面发挥作用,同时可能作为一种相关因子影响关键酶活性,改变细胞壁多糖网状结构以至细胞壁的延展性[9]。近年研究表明培养基中添加一定浓度硼酸有利于花粉萌发生长[10-12]。杜纪红等[13]认为不同植物种类的花粉萌发和花粉管生长所需的最适硼酸浓度不同:如培养黄薇花粉适宜硼酸浓度为 500~1000 mg/L[14];150 mg/L 硼酸能显著提高福鼎大白茶花粉萌发率[15];王玉林等[16]发现添加 100 mg/L 硼酸可使火龙果花粉离体萌发率显著提高 59.75%,而花粉管生长需要 500 mg/L 高浓度的硼酸;因此有必要对不同的植物开展研究。本研究结果表明,多肉植物红司花粉萌发适宜的硼酸为 60~100 mg/L,而花粉管的伸长仅需要 30 mg/L 低浓度的硼酸,随着浓度的增加,花粉管生长受到抑制。
除了硼酸之外,温度是影响萌发率及花粉管生长的重要因素,本实验设置的处理温度范围为 12~44 ℃,从低温区覆盖至高温区,数据分析结果显示不同水平的温度处理对花粉萌发和花粉管生长影响差异极显著(P<0.01)。不同植物种类适宜花粉萌发的温度有差异。地被菊的花粉适宜萌发温度只有 10~15 ℃[17],梅花等早春开花植物花粉萌发最佳温度 20 ℃左右[18],茉莉花粉最佳培养温度为 25~30 ℃[19],即使种内不同品种的花粉适宜萌发温度也有较大差异,实验室前期研究发现,拟石莲属一些早开花品种如‘花月夜’‘灵隐’ 的花粉在 15 ℃低温离体培养下花粉活力较高,平均萌发率分别达到 66.67%、29.51%。而品种‘莎维娜’的花粉在 30 ℃高温培养下花粉萌发率达到 26.67%。因此有必要针对重要的育种材料开展研究。红司花粉萌发的适宜温度范围较广,试验表明 12~28 ℃温度下花粉萌发率均较高,其中 12 ℃ 低温条件下花粉萌发效果最佳(21.86%);12 ℃ 是本试验设置的低温,如果温度更低,萌发率有可能更高,因此可以在实际育种工作中,尝试更低温度处理红司花粉。相关性分析发现温度与萌发率呈负相关,与花粉管长度呈正相关,说明花粉萌发和花粉管生长可能需要不同的温度条件。本试验中,12~28 ℃培养有利于提高花粉管萌发率,20~36 ℃有利于花粉管的伸长,与蒿柳研究结果类似[20]。这对多肉植物育种可能有很好的指导作用,可以尝试前期低温诱导花粉大量萌发后,提高培养温度促进花粉管伸长。
不同植物种类花粉管生长适宜观察的培养时间存在差异。海仙花花粉培养 6 h 花粉管生长最快,状态最好[21];矾根‘红妆魅影’培养时间达到 2 h 时,矾根花粉管发生明显伸长[22];野杏花粉离体培养最佳时间为 6 h[23]。本研究为了探明红司花粉在 24 h 内萌发及花粉管生长情况,同时基于试验的可操作性。从早上 9 点取新鲜花粉处理后,放在不同处理下分别培养 1、3、16、24 h,取出观察拍片。数据分析结果表明:时间是影响红司花粉萌发率的极显著因素(P<0.01),对花粉管长度影响则不显著(P>0.05)。空白对照中红司花粉有 3.71% 少量萌发,平均花粉长度为 44.40 μm。处理 1 h 后花粉的平均萌发率升高至 10.43%,花粉管长度为 130.42 μm。随着培养时间的延长,3~16 h 花粉萌发率迅速提高,花粉管生长加快。培养至 16 h,平均萌发率(18.70%)与花粉管长度(194.33 μm)均达到最大值。培养时间的增加有利于花粉萌发率的上升及花粉管的伸长,但时间过长,部分已萌发的花粉出现花粉管破裂,扭曲交缠,不利于数据统计,这也是花粉在培养 24 h 之后萌发率在数据上出现下降原因之一。综上所述,红司花粉离体培养适宜的观察时间可能在 3~16 h 之间,需要在实际育种工作中进一步缩小变量范围,摸索最佳观察时间。
方差分析显示,硼酸浓度、培养温度、培养时间及交互效应对花粉萌发率的影响差异极显著,60 mg/L 硼酸+12 ℃+16 h 培养下,萌发率达到最大值 52.43%,极大的提高了花粉萌发率。硼酸浓度、培养温度及交互效应对花粉管伸长的影响极显著。培养基中添加 30 mg/L 硼酸 36 ℃培养 3~24 h 后,400~1100 μm 长度的花粉管比例最高,平均数量为 35 个,平均花粉管长度为 3 h: 453.16 μm、16 h:527.74 μm、24 h:571.52 μm,培养 24 h 虽然花粉管平均长度值最大,但 400 μm 以内的花粉管数量仅为 5 个,不利于提高授粉成功率。综合分析认为 60 mg/L 硼酸+12 ℃+16 h (A3B1C3)为红司花粉萌发较理想条件,30 mg/L 硼酸+36 ℃+16 h(A2B4C3)为红司花粉管生长的适宜条件。——论文作者:杨 澜1,2,张朝君1,2,杜致辉1 ,陈之林1*
参考文献
[1] González-Mancera G, Reyes-Santiago J, Luna-Islas M A, et al. Morphological characterization of pollen from some species of the genus Echeveria. from Mexican territory[J]. Microscopy and Microanalysis, 2016, 22(S3): 1048-1049.
[2] Rodríguez-Rojas T J, Andrade-Rodríguez M, Canul-Ku J, et al. Pollen viability, stigma receptivity and pollination type in five Echeveria species under greenhouse conditions[J]. Revista Mexicana de Ciencias Agrícolas, 2015, 6(1): 113-127.
[3] Jimeno-Sevilla H D, Hernández-Ramírez A M, Ornelas J F, et al. Morphological and nectar traits in Echeveria rosea Lindley (Crassulaceae) linked to hummingbird pollination in central Veracruz, Mexico[J]. Haseltonia, 2014, 19: 17-25.
[4] Parra-Tabla V, Vargas C F, Eguiarte L E. Is Echeveria gibbiflora (Crassulaceae) fecundity limited by pollen availability? An experimental study[J]. Functional Ecology, 1998, 12(4): 591-595.
[5] 谢深喜, 罗先实, 吴月嫦, 等. GA3、2,4-D、B 和蔗糖对梨花粉生活力及花粉生长速度的影响[J].果树学报, 2004, 21(4): 289-294.
[6] 陈 奇, 张树娟, 孙万仓, 等. 白菜型油菜花粉离体萌发液体培养基组分的优化[J]. 分子植物育种, 2017, 15(7): 2742-2748.
[7] Stenli R G, Linskens H F. Pollen: Biology biochemistry management [M]. Berlin Heidelberg: Springer-Verlag, 1974: 37.
[8] 张 琛, 裴嘉博, 郗笃隽, 等. 甜樱桃花粉萌发率测定条件的优化研究[J]. 落叶果树, 2018, 50(6): 15-17.
[9] 杨晓东, 孙素琴, 李一勤. 硼缺乏导致花粉管细胞壁多糖分布的改变[J]. 植物学报, 1999, 41(11): 1169.
[10] 韩 霜. 钙和硼对菊花品种金陵丰收花粉萌发及花粉管生长的影响[J]. 江苏农业科学, 2016, 45(5): 258-260, 261.
[11] 毛立彦, 唐毓玮, 谢振兴, 等. ‘保罗兰’睡莲花粉离体萌发及花粉管生长的研究[J]. 西南农业学报, 2019, 32(5): 1155-1161.
[12] 郭英姿, 贾文庆, 周秀梅, 等. 早开堇菜花粉生活力研究[J]. 江苏农业科学, 2019, 47(13): 191-194.
[13] 杜纪红, 叶正文, 苏明申, 等. 桃花粉离体萌发和花粉管生长特性研究[J]. 西北植物学报, 2011, 31(1): 64-71.
[14] 罗卿清, 郑 钢, 顾翠花, 等. 黄薇花粉活力及柱头可授性[J]. 浙江农林大学学报, 2020, 37(1): 182-187.
[15] 林开勤, 刘声传, 梁思慧, 等. 茶树花粉离体萌发条件优化及活力快速检测[J]. 种子, 2018, 37(12): 14-18.
[16] 王玉林, 胡位荣, 刘顺枝, 等. 硼酸、蔗糖和 pH 值对火龙果花粉离体萌发和花粉管伸长的影响[J]. 分子植物育种, 2018, 16(1): 240-247.
