
 >
>稀有种和常见种对黄土高原辽东栎群落物种多样性贡献的多尺度分析
时间:
摘要: 明确物种多样性的多尺度格局及其形成过程是进行物种多样性保护的重要前提。通过加性分配的方法,在 α1( 小样方内) 、β1( 小样方间) 、β2( 样方间) 、β3( 样地间) 4 个空间尺度上,研究了黄土高原子午岭辽东栎( Quercus wutaishanica) 群落植物多样性的多尺度变化,确定了稀有种( 频度<5%) 和常见种( 频度>25%) 对物种多样性格局的相对贡献及其尺度依赖性,旨在阐明黄土高原辽东栎群落物种多样性的多尺度格局及其形成过程,为该区物种多样性的保护提供理论依据。结果表明: ( 1) 乔、灌、草 3 层呈现了相似的物种多样性格局。物种丰富度基本上呈现了随尺度的增大而增加的变化趋势,小样方尺度上的α 多样性( α1) 对群落物种丰富度的贡献最小,样地尺度上的 β 多样性( β3) 对各层物种丰富度的贡献最大( β3 > 50%) 。( 2) Shannon 多样性呈现了与物种丰富度截然相反的格局,小样方尺度上的 α 多样性成分( α1) 是 Shannon 多样性的主要贡献者( α1 > 45%) ,尔后随尺度的增大,Shannon 多样性逐渐减小。( 3) 去除常见种引起的物种丰富度减小效应表现为随尺度的增加,影响逐渐减小,其效应主要集中在小样方内( α1 尺度) 以及小样方间( β1 尺度) 2 个尺度上; 而去除稀有种引起的物种丰富度减小效应表现为随尺度的增加,影响逐渐增大,其效应主要集中在样方间( β2) 以及样地间( β3) 2 个尺度上。( 4) 去除稀有种对 Shannon 多样性的影响在每一尺度上均不明显,去除前后的曲线几乎重合,而去除常见种对 Shannon 多样性的影响在每一尺度上均十分明显。其中,去除常见种,草本层 Shannon 多样性在每一尺度都相应增大,而对于灌木层和乔木层,Shannon 多样性则表现为在 α1 尺度上减小,而在其他尺度上增大。本研究揭示了空间尺度在评价稀有种和常见种对植物群落物种多样性相对贡献中的重要性。稀有种主要在较大尺度( β2 和 β3) 上影响物种丰富度格局,而常见种不仅在小尺度( α1 和 β1) 上影响物种丰富度,而且在较大尺度( β2 和 β3) 上影响物种均匀度。同时,物种多样性的加性分配也是在多尺度上揭示物种多样性变化格局以及变异来源的有效且比较简单的方法。
关键词: 空间尺度; 加性分配; β 多样性; 稀有种; 常见种
明确物种多样性格局及其形成过程是进行物种多样性保护的重要前提[1-2]。近年来,物种多样性的很多研究都集中在探索决定物种多样性空间格局的生态过程,如扩散限制[3]、环境筛选[4]以及二者相对作用的分解等方面[5-7]。事实上,群落中不同的物种类群组成也是决定物种多样性格局的重要因素,研究不同物种类群组成对物种多样性空间格局的相对贡献也是探索物种多样性格局形成过程的另外一个重要途径[8-10]。
近年来,学者们已经开展了大量有关物种类群组成,如不同的功能群组成[11]、不同的稀有种和常见种比例等[12]对物种多样性空间格局影响的研究,诚然,稀有种( 狭布种) 和常见种( 广布种) 对物种多样性的相对贡献争议 最 大[12-13]。大 量 研 究 表 明,常见种多样性与群落总体物种多样性的相关性要明显高于稀有种[10,13-15]。将群落总体物种多样性进一步分解为 α 多样性和 β 多样两种成分,研究发现,常见种对 α 多样性和 β 多样性的贡献也均明显大于稀有种[15]。因此,很多研究认为,常见种是决定物种多样性格局的主要成分,这与人们的长期认识完全不同,即物种多样性应该由大量的分布受限的稀有种决定,而不是由数量不多但是分布范围广且多度大的常见种决定[16]。关于常见种为什么不对称地决定群落总体物种多样性格局仍没有很好的解释,一个直接的证据是常见种与群落环境的相关性明显高于稀有种,但也仅此而已[7,17-18]。也有研究表明,常见种对物种多样性较高的贡献率可能更多的是由于有偏差的取样效应引起的,而非真正的稀有种和常见种的生物学差异所致[19]。但是,这些结论都是基于单一尺度上的研究,而物种多样性格局的形成过程是尺度依赖性的,即存在尺度效应。因此,在多尺度上研究稀有种和常见种对物种多样性的相对贡献可能是解决争议的重要途径。目前,有关物种多样性格局的多尺度研究[20-24]以及稀有种和常见种对物种多样性相对贡献的研究相对较多[10,18],而关于稀有种和常见种对物种多样性相对贡献的多尺度分析鲜有报道。
辽东栎( Quercus wutaishanica) 隶属于壳斗科( Fagaceae) 栎属( Quercus) ,是我国暖温带极为重要的森林建群种,也是我国暖温带落叶阔叶林的主要优势种和华北针阔混交林的主要成分[25]。目前,关于辽东栎群落物种多样性的研究主要集中在物种多样性空间格局的研究方面[25-28],而缺少对决定物种多样性格局的生态过程的研究,特别是缺少多尺度上的研究。β 多样性是度量生态梯度上的物种周转( directional turnover along a gradient) 和非梯度上的物种组成变异( nondirectional variation) 的重要参数,其有效连结了局域尺度上的 α 多样性和区域尺度上的 γ 多样性。关于 β 多样性的测量指标有很多[29],本研究通过物种多样性加性分配方法来构建 α、β、γ 三者之间的关系( γ =α + β) 。在加性分配模型中,α、β 和 γ 多样性都具有相同的表达方式和量纲,可以进行直接比较,能在多个尺度上解释 α 和 β 多样性对区域总的 γ 多样性的相对贡献[30]。
辽东栎是黄土高原子午岭林区气候顶极群落的建群种,以辽东栎为优势种的森林群落对区域内的物种多样性的恢复与保护具有重要作用。本研究以位于黄土高原中部子午岭的天然辽东栎群落为研究对象,通过物种多样性的加性分配方式在多尺度上确定: ( 1) α 和 β 多样性对区域上总体 γ 多样性的相对贡献及其尺度依赖性; ( 2) 通过去除稀有种和常见种引起的物种多样性变化,确定稀有种和常见种对物种多样性格局的相对贡献及其尺度依赖性,旨在阐明辽东栎群落物种多样性的多尺度格局及其形成过程,为该区物种多样性的保护提供理论依据。
1 研究方法
1.1 样地设置
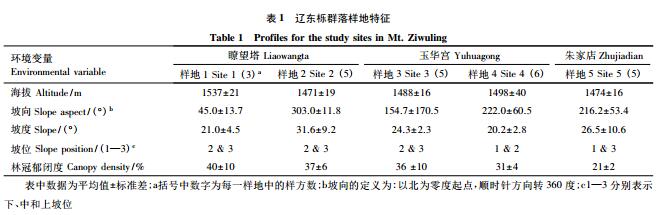
在对黄土高原中部子午岭全面考察的基础上,选择具有代表性的辽东栎林进行分层抽样调查,形成小样方-样方-样地的 3 级取样,共设置 5 个样地。其中,瞭望塔( 35°16' N,108°36' E) 2 个样地,朱家店( 35°19' N, 108°41' E) 1 个样地以及玉华宫( 35°19' N,108°53' E) 2 个样地。为消除边缘效应,取样样方尽可能地设置在样地中心位置,在瞭望塔的 2 个样地分别设置 3 个和 5 个样方,在玉华宫的 2 个样地分别设置 6 个和 5 个样方,在朱家店样地设置 5 个样方,总共建立了 24 个 20 m×20 m 的样方。目前的研究中,群落中的物种多样性是按照生长型分层调查的,但这样的不足是,忽略了物种本身对环境等其他生态过程的响应,因此,基于植物的形态、生理、生活史或其他相关的生态系统过程以及与物种相联系的某些生物学特征,将群落中的物种划分为不同的功能群更加合理[31]。本研究中,样方中的物种根据其生长型和分布的垂直结构,划分成乔木层( 高度>3 m) 、灌木层( 高度介于 1 m 和 3 m 之间) 和草本层( 高度<1 m) 3 层作为功能群单元分别调查。每个样方分成 4 个 10 m×10 m 的小样方进行乔木层物种调查,在样方每个象限的中心分别设置 1 个 4 m×4 m 和 1 个 1 m×1 m 的小样方进行灌木层和草本层物种调查,每层共计 96 个小样方。在每个小样方记录物种种类、盖度、数量、高度等数量指标,同时记录样方的经纬度、海拔、坡度、坡向、坡位以及林冠郁闭度等环境特征( 表 1) 。
1.2 物种多样性的多尺度分析
根据物种多样性加性分配框架[20,32],区域上物种多样性在小样方、样方和样地 3 取样个尺度上的分配关系为( 表 2) : γ( 区域尺度上总物种多样性,total species diversity) = α1( 小样方内物种多样性,within subplot) +β1( 小样方间物种多样性,between subplots) + β2( 样方间物种多样性,between plots) + β3( 样地间物种多样性,between sites) 。
由于物种丰富度与生态系统的功能密切相关,常常被认为是度量物种多样性的代表性指数[33]。同样的,物种均匀度也可以影响群落的构建过程以及生态系统功能[34]。因此,本研究选取物种丰富度和 Shannon 多样性 2 种多样性指数进行研究,其中,物种丰富度以物种种类为测量特征,而 Shannon 多样性指数是反映群落物种丰富度和均匀度的综合指标。
1.3 稀有种和常见种对物种多样性的相对贡献
为了研究稀有种和常见种对物种多样性的相对贡献,参照相关文献[35]和子午岭辽东栎群落物种分布的实际情况,本研究采用频度定义物种的稀有性和常见性,其中,96 个小样方中频度<5%的物种为稀有种,频度>25%的为常见种。在 α1( 小样方内) 、β1( 小样方间) 、β2( 样方间) 、β3( 样地间) 4 个尺度上,通过比较物种去除前后所引起的多样性变化程度来确定该类物种( 即稀有种和常见种) 对物种多样性格局的相对贡献。
2 结果
在 96 个小样方中,记录到草本层植物 80 种,灌木层植物 102 种和乔木层植物 34 种,共计 216 个物种。其中,稀有种( 频度<5%) 131 种,占到各层物种比例的 53%—71%; 常见种( 频度>25%) 32 种,占到各层物种比例的 4%—26%。小样方中草本层的平均物种数为 4.68,灌木层为 12.26,乔木层为 4.59( 表 3) 。
2.1 辽东栎群落物种多样性的多尺度格局
乔、灌、草 3 层物种丰富度的加性分配均呈现了相似的多样性格局。物种丰富度基本上呈现了随尺度的增大而增加的变化趋势,其中,小样方尺度上的 α 多样性( α1) 对群落物种丰富度的贡献最小,样地尺度上的 β 多样性贡献最大( β3>50%) ( 图 1) 。
Shannon 多样性的加性分配呈现了与物种丰富度截然相反的多样性格局。总体上,乔、灌、草 3 层都呈现了较为相似的变化趋势,即随尺度的增大,Shannon 多样性逐渐减小( 图 2) 。在物种丰富度格局中,小样方尺度上的 α 多样性( α1) 对总体物种多样性的贡献<15%,而其对 Shannon 多样性的贡献则>45%,小样方尺度上的 α 多样性成分( α1) 是 Shannon 多样性的最大贡献者( 图 2) 。需要注意的是,Shannon 多样性在不同层次间的变化也不尽相同,三层中灌木层在各尺度上均具有较草本层和乔木层较大的 Shannon 多样性,同时,草本层的 β3 尺度上也具有较大的贡献值( 图 2) 。
稀有种和常见种对黄土高原辽东栎群落物种多样性贡献的多尺度分析推荐阅读:高级农业经济师论文要求
2.2 稀有种和常见种对物种多样性的相对贡献
稀有种和常见种对物种丰富度的影响呈现了明显不同的变化趋势。去除常见种引起的物种丰富度减小效应表现为随尺度的增加,影响逐渐减小,其效应主要集中在小样方内( α1) 以及小样方间( β1) 2 个尺度上。而稀有种则刚好相反,去除稀有种引起的物种丰富度减小效应表现为随尺度的增加,影响逐渐增大,其效应主要集中在样方( β2) 以及样地( β3) 2 个尺度( 图 3) 。
稀有种和常见种对群落物种多样性的影响不仅表现在去除二者成分引起的物种多样性变化的尺度依赖性,还表现在二者对不同物种多样性指数的依赖性。与物种丰富度不同,去除稀有种对 Shannon 多样性的影响在每一尺度上均不明显,去除前后的曲线几乎重合,而去除常见种对 Shannon 多样性的影响在每一尺度上均十分明显( 图 4) 。其中,去除常见种,草本层 Shannon 多样性在每一尺度都相应增大,而对于灌木层和乔木层,Shannon 多样性则表现为在 α1 尺度上减小,而在其他尺度上增大。
3 讨论
无论局域还是区域上都常常拥有大量的稀有种,却仅有较少的常见种,但是,越来越多的研究已经表明,常见种才是决定群落物种多样性格局的重要类群,这是一个违背直觉的事实[36]。对黄土高原辽东栎群落物种多样性的单一取样尺度上的研究表明,常见种不仅主导群落的总体物种丰富度格局,同时也是 α 多样性和 β 多样性格局的重要贡献者。因此,常见种被认为是群落物种丰富度格局的指示者,也应该是优先保护的物种[28]。然而,本研究在 α1( 小样方内) 、β1( 小样方间) 、β2( 样方间) 、β3( 样地间) 4 个尺度上进行了物种多样性分析,结果表明,常见种的确是物种多样性的重要贡献者,但决不能忽略稀有种在大尺度上对物种丰富度的重要贡献,因为稀有种和常见种对物种多样性的贡献是尺度依赖的。在物种多样性保护中应注意空间尺度的重要性,物种多样性的加性分配方法是有效洞察物种多样性多尺度格局的有效途径[20,32,37-38]。
物种丰富度加性分配的结果表明,乔、灌、草 3 层均呈现了相似的物种多样性格局。物种丰富度随尺度的增加而增加,样地尺度上的 β 多样性( β3) 对区域上总体物种多样性的贡献最大( 图 1) 。这与很多群落物种多样性的多尺度格局相似,即物种丰富度的较大贡献者常常来源于大尺度上较大的 β 多样性[20,32,39]。例如, Chandy 等[39]的研究表明,对乔木层和林下木本层的物种多样性来说,较大的物种多样性贡献也来自于较大尺度( 样方尺度和研究自然区域尺度) 的 β 多样性。同样的,对北京东灵山辽东栎群落物种多样性的多尺度研究也得到了相似的结果[21]。这些结果均暗示 β 多样性在区域物种多样性保护中的重要性。β 多样性是指取样单元间物种多样性的差异或者转换,因此,保持不同取样单元间的异质性将会极大地保护区域上的物种多样性。在本研究中,辽东栎群落的物种丰富度随取样尺度( α1、β1、β2、β3) 的增加而增加( 图 1) ,这主要是因为黄土高原子午岭辽东栎林中的多数物种是局域分布的,少数物种是区域分布的,在多数样方内都是少数相同的常见种组成,而绝大多数偶见种都仅仅在少数样方中出现,这就导致了取样单元间较高异质性( 即物种组成的不相似性) ,并且这种异质性随尺度的增加而增加。因此,物种丰富度主要由稀有种的差异引起,且主要在较大尺度上( 如 β2、β3) 起作用( 图 3; 图 5) 。在黄土高原,由于战争等历史原因,加之近年来人类活动不断促进了新斑块的产生[40],天然辽东栎群落呈斑块状分布[41]。就植物种类本身而言,生境破碎化对群落中稀有种的影响更大,由于该区较严重的生境破碎化[42],这直接限制了斑块间的物种扩散,增加了这些群落中稀有种和特有种的数量[43],从而决定了区域尺度上较大的 β 多样性。很多研究已经表明,β 多样性是大多数生态系统物种多样性维持的重要成分[44],同时,β 多样性在物种多样性格局中的重要性与生境破碎化程度有关[43],应该引起物种多样性保护工作者的足够重视。
